



Hereditary Colorectal Cancer and Polyposis Panel
Summary and Pricing 
Test Method
Sequencing and CNV Detection via NextGen Sequencing using PG-Select Capture Probes| Test Code | Test Copy Genes | Panel CPT Code | Gene CPT Codes Copy CPT Code | Base Price | |
|---|---|---|---|---|---|
| 5467 | Genes x (25) | 81435 | 81201(x1), 81203(x1), 81292(x1), 81294(x1), 81295(x1), 81297(x1), 81298(x1), 81300(x1), 81317(x1), 81319(x1), 81321(x1), 81323(x1), 81403(x1), 81404(x1), 81405(x3), 81406(x3), 81408(x1), 81479(x29) | $990 | Order Options and Pricing |
Pricing Comments
Testing run on PG-select capture probes includes CNV analysis for the gene(s) on the panel but does not permit the optional add on of exome-wide CNV analysis. Any of the NGS platforms allow reflex to other clinically relevant genes, up to whole exome or whole genome sequencing depending upon the base platform selected for the initial test.
An additional 25% charge will be applied to STAT orders. STAT orders are prioritized throughout the testing process.
This test is also offered via a custom panel (click here) on our exome or genome backbone which permits the optional add on of exome-wide CNV or genome-wide SV analysis.
Turnaround Time
3 weeks on average for standard orders or 2 weeks on average for STAT orders.
Please note: Once the testing process begins, an Estimated Report Date (ERD) range will be displayed in the portal. This is the most accurate prediction of when your report will be complete and may differ from the average TAT published on our website. About 85% of our tests will be reported within or before the ERD range. We will notify you of significant delays or holds which will impact the ERD. Learn more about turnaround times here.
Targeted Testing
For ordering sequencing of targeted known variants, go to our Targeted Variants page.
Clinical Features and Genetics
Clinical Features
Colorectal cancer (CRC) is the development of tumors in the colon and rectum that occur in approximately 8% of individuals with cancer (Siegel et al. 2015. PubMed ID: 25559415). Similar to other cancers, risk for CRC depends on both genetic and environmental factors (smoking, diet). Approximately 30% of CRCs are considered familial, and 5% of CRCs are caused by a Mendelian disorder (Esteban-Jurado et al. 2014. PubMed ID: 24587672). Colorectal cancer is generally broken down into the presence or absence of polyposis (numerous internal polyps). Identification of pathogenic variants in the germline of CRC patients is important for cancer surveillance (colonoscopy) for the affected individual and family members, since early detection and treatment have been shown to decrease morbidity and mortality (Kohlmann and Gruber. 2018. PubMed ID: 20301390). Different types of CRC inherited diseases are described below.
Lynch Syndrome: Lynch syndrome, also known as hereditary nonpolyposis colorectal cancer (HNPCC), is an inherited cancer syndrome mainly caused by germline pathogenic variants in DNA mismatch repair (MMR) genes. MMR genes are responsible for repairing small sequence errors, or mismatches, during DNA replication. Pathogenic variants in mismatch repair genes can cause widespread genomic instability characterized by the expansion and contraction of short tandem repeat sequences (microsatellites; Grady and Carethers. 2008. PubMed ID: 18773902). As a result, Lynch syndrome is marked by early onset and a high lifetime risk of cancers, particularly in the right colon but also in the endometrium, ovary, stomach, bile duct, kidney, bladder, ureter, and brain (Jang and Chung. 2010. PubMed ID: 20559516).
Hereditary Diffuse Gastric Cancer (HDGC): Hereditary diffuse gastric cancer is a highly penetrant, diffuse-type gastric cancer. Patients with HDGC typically present at about 40 years of age and have a cumulative cancer risk of 67% for men and 83% for women by 80 years of age (Pharoah et al. 2001. PubMed ID:11729114). HDGC is caused by CDH1 pathogenic variants, which are found in all ethnic groups but are rare in countries with high rates of sporadic gastric cancer, such as Japan and Korea (Guilford et al. 2010. PubMed ID: 20373070). Approximately 10% of gastric cancers show familial clustering, and about 1–3% of cases are known to be hereditary (e.g., CDH1 pathogenic variants). Pathogenic variants in CDH1 can also cause lobular breast cancer in women (Carneiro et al. 2008. PubMed ID: 17513507).
Juvenile Polyposis Syndrome (JPS): Juvenile polyposis syndrome is a rare, inherited hamartomatous polyposis syndrome with increased susceptibility to colorectal cancer. Clinical diagnosis of JPS is typically made when one of the following criteria is met: more than five juvenile polyps in the colorectum; multiple juvenile polyps throughout the GI tract; or any number of juvenile polyps and a family history of gastrointestinal polyps (Chow and Macrae. 2005. PubMed ID: 16246179). Juvenile refers to the developmentally immature nature of the polyp, not the age of disease onset. In addition to polyposis, 10-20% of JPS patients also have extracolonic abnormalities such as congenital heart defects, cleft lip or palate, microcephaly, and malrotations (Eng et al. 2001. PubMed ID: 11160785). Even though a solitary juvenile polyp in the general population has very little malignant potential (Nugent et al. 1993. PubMed ID: 8395444), patients with JPS have a 68% chance of developing gastrointestinal cancer by the age of 60 (Chow and Macrae. 2005. PubMed ID: 16246179). Thus, confirming a diagnosis of JPS is important for the appropriate surveillance and management of cancer.
PTEN Hamartoma Syndrome: PTEN hamartoma tumor syndrome (PHTS) is a cluster of related clinical conditions, all caused by germline pathogenic variants in the PTEN tumor suppressor gene. Included in PHTS are Cowden syndrome (CS), Bannayan-Riley-Ruvalcaba syndrome (BRRS), Proteus syndrome (PS) and Proteus-like syndromes, and VACTERL association with hydrocephalus. While each PHTS condition has its own unique pathognomonic features (Blumenthal and Dennis. 2008. PubMed ID: 18781191), hamartomatous overgrowth, macrocephaly, and vascular malformations appear to be common to all conditions (Zhou et al. 2001. PubMed ID: 11476841). A presumptive diagnosis of PHTS is typically made based on clinical symptoms, but a definitive diagnosis requires the identification of a heterozygous PTEN pathogenic variant. Patients with a germline pathogenic variant in PTEN have a 5- to 10-fold higher chance of developing cancer at a much earlier age (<30 years) than the general population (Eng. 2003. PubMed ID: 12938083). In addition to confirming the diagnosis of PHTS, testing patients for a germline PTEN pathogenic variant is essential to accurately assess their risk for cancer and to make appropriate recommendations regarding prevention and treatment of malignancy.
Familial Adenomatous Polyposis (FAP): Familial adenomatous polyposis is an inherited cancer syndrome characterized clinically by the development of hundreds to thousands of adenomatous polyps in the colon and rectum. If untreated, nearly all FAP patients will develop CRC by age 40 (Fearnhead et al. 2001. PubMed ID: 11257105). In addition to CRC, FAP patients are also predisposed to desmoid tumors, small bowel cancer, thyroid cancer, hepatoblastoma, and medulloblastoma (Galiatsatos and Foulkes. 2006. PubMed ID: 16454848). About 60% of families with FAP also display congenital hypertrophy of the retinal pigment epithelium (CHRPE), a condition that does not affect sight or have malignant potential but can be easily detected by ophthalmoscopy at any age. CHRPE is highly diagnostic of FAP and can be useful for identifying FAP patients and at-risk family members, well before the appearance of polyps (Díaz-Llopis and Menezo. 1988. PubMed ID: 2830869).
Peutz-Jeghers Syndrome (PJS): Peutz-Jeghers syndrome is characterized by hamartomatous polyps in the gastrointestinal tract and melanin pigmentation around the mouth, eyes, nostrils, buccal mucosa, fingers, toes, and other sites. PJS patients typically present in early childhood with abnormal pigmentation or with complications of polyposis, such as intussusception, bowel obstruction, or bleeding. Compared to the general population, patients with PJS have an increased risk of intestinal and various extra-intestinal malignancies, including breast, pancreatic, ovarian, testicular, and cervical cancer; their lifetime risk is ~4-fold higher for gastrointestinal cancer and ~6-fold higher for breast cancer compared to individuals without PJS (Hearle et al. 2006. PubMed ID: 16707622). Approximately 75% of PJS cases are known to be familial; the remainder appears to be sporadic (Lim et al. 2003. PubMed ID: 12865922).
Li-Fraumeni Syndrome (LFS): Li-Fraumeni syndrome is a hereditary cancer syndrome that predisposes individuals to multiple neoplasms at an early age. The most common neoplasms associated with LFS are bone and soft-tissue sarcomas, pre-menopausal breast carcinomas, adrenocortical carcinomas, and brain tumors. Although much less common, melanomas, germ cell tumors, gastric carcinomas, and Wilms tumors have also been described in LFS patients (Varley et al. 1997. PubMed ID: 9242456). The average age of malignancy for individuals with LFS is typically between 20 and 45, which is at least 2-3 decades sooner than reported for the general population (Nichols et al. 2001. PubMed ID: 11219776).
CHEK2-related Cancers: Pathogenic variants in the CHEK2 gene have been reported to cause a Li-Fraumeni-like syndrome (Bell et al. 1999. PubMed ID: 10617473), although subsequent studies have indicated that CHEK2 pathogenic variants are only very rarely found in patients with classic symptoms of LFS (Lee et al. 2001. PubMed ID: 11719428). Pathogenic variants in CHEK2 have been more frequently found in patients who have hereditary breast cancer (Vahteristo et al. 2002. PubMed ID: 12094328; Meijers-Heijboer et al. 2003. PubMed ID: 12690581). CHEK2 pathogenic variants have also been reported in a family with breast and colon cancer, and specific variants may predispose individuals to colorectal cancer (Abud et al. 2012. PubMed ID: 23329222; Narod. 2010. PubMed ID: 20597917).
MUTYH-Associated Polyposis (MAP): MUTYH-associated polyposis is an autosomal recessive form of FAP caused exclusively by pathogenic variants in the MUTYH gene (Al-Tassan et al. 2002. PubMed ID: 11818965; Sieber et al. 2003. PubMed ID: 12606733). Individuals with MAP typically present by age 55 with multiple (between 10 and 1,000) colorectal adenomas, some of which have or will become colorectal tumors (Poulsen and Bisgaard. 2008. PubMed ID: 19506731).
Ataxia-Telangiectasia: Ataxia-telangiectasia (A-T) is characterized by early-onset (1-4 years) progressive cerebellar ataxia, telangiectasias of the conjunctivae, oculomotor apraxia, choreoathetosis, immunodeficiency, slurred speech, frequent infections, and an increased risk of cancers, especially leukemia and lymphoma. Unlike A-T, which is caused by homozygous or compound heterozygous pathogenic variants in the ATM gene, heterozygous carriers of an ATM causative variant are at an increased risk of breast cancer (Concannon et al. 2008. PubMed ID: 18701470) and heart disease (Swift et al. 1991. PubMed ID: 1961222). In addition, one mutated copy of ATM has been associated with an increased risk of stomach and colorectal cancer (Thompson et al. 2005. PubMed ID: 15928302).
Oligodontia-Colorectal Cancer: Oligodontia is a congenital condition where six or more permanent teeth fail to develop (Matalova et al. 2008. PubMed ID: 18573979; De Coster et al. 2009. PubMed ID: 18771513). Pathogenic variants in the AXIN2 gene have been associated with oligodontia and colorectal cancer (Lammi et al. 2004. PubMed ID: 15042511). In patients with AXIN2 pathogenic variants, oligodontia presents at an early age when their permanent teeth fail to develop, whereas colorectal neoplasia develops much later, usually between the ages of 30 and 60. Accordingly, screening patients with non-syndromic oligodontia for mutations in the AXIN2 gene can be an effective way to identify individuals with a risk of developing colorectal cancer later in life.
Genetics
The colorectal cancer next generation sequencing panel assesses genes that have been shown to be causative when mutated for disorders that have colorectal cancer as a clinical feature.
MLH1, MSH2, MSH6, PMS2, and EPCAM: Lynch syndrome is an autosomal dominant disease mainly caused by germline pathogenic variants in one of four MMR genes: MLH1, MSH2, MSH6, and PMS2 (Peltomäki and Vasen. 2004. PubMed ID: 15528792; Kohlmann and Gruber. 2018. PubMed ID: 20301390). Pathogenic variants in the MLH1 and MSH2 genes account for 80-90% of all Lynch syndrome patients and most frequently occur in families meeting the stringent Amsterdam I criteria. Pathogenic variants in the MSH6 and PMS2 genes account for most of the remaining Lynch patients and are often found in families with atypical symptoms, such as extracolonic carcinomas; these patients have also been found to have a low rate of microsatellite instability.
Pathogenic variants in another gene, EPCAM, which encodes a calcium-independent cell adhesion molecule and not a mismatch repair protein, are also involved in Lynch syndrome. Germline pathogenic variants in the EPCAM gene cause inactivation of the nearby MSH2 gene via hypermethylation in approximately 1-3% of individuals with Lynch syndrome (Kohlmann and Gruber. 2018. PubMed ID: 20301390). The only reported pathogenic variants of the EPCAM gene that are causative for Lynch Syndrome are large deletions. Missense, nonsense, and splicing pathogenic variants are involved in congenital tufting enteropathy (Human Gene Mutation Database; www.insight-group.org). The cumulative incidence of colon cancer risk from EPCAM deletions has been estimated to be 75% by 70 years of age, and 12% for endometrial cancer in women (Kempers et al. 2011. PubMed ID: 21145788).
A germline inversion of exons 1-7 in MSH2 has been reported in fourteen individuals from eleven unrelated families who clinically presented with Lynch syndrome-associated phenotypes including colorectal, endometrial, gastric, and ovarian cancer (Wagner et al. 2002. PubMed ID: 12203789; Rhees et al. 2013. PubMed ID: 24114314; Mork et al. 2016. PubMed ID: 28004223).
CDH1: Hereditary diffuse gastric cancer (HDGC) is an autosomal dominant disease that is caused by pathogenic variants in the CDH1 gene. CDH1 encodes epithelial cadherin (E-cadherin), which is a transmembrane protein responsible for cell-to-cell adhesion and cellular invasion suppression. It also plays important roles in signal transduction, differentiation, gene expression, cell motility, and inflammation (Kaurah and Huntsman. 2014. PubMed ID: 20301318). The activity of E-cadherin in coordination with the actin cytoskeleton through catenins (α-, β-, and γ-) is responsible for cellular adhesion. Many human cancers show low levels of E-cadherin compared to normal tissue, which causes defects in cellular adhesion and ultimately leads to metastasis. HDGC is mostly inherited from an affected family member. The majority of germline pathogenic variants are truncating mutations in E-cadherin; the second hit of the normal allele in gastric tissue is usually due to CDH1 promoter hypermethylation and secondly to CDH1 deletions (Schrader and Huntsman. 2010. PubMed ID: 20517687).
BMPR1A and SMAD4: Juvenile polyposis syndrome (JPS) is caused by heterozygous germline pathogenic variants in one of two genes: BMPR1A and SMAD4 (Howe. 1998. PubMed ID: 9582123; Howe et al. 2001. PubMed ID: 11381269). Both genes mediate the biological effects of the transforming growth factor-β (TGF-β) superfamily of cytokines (Miyazono et al. 2009. PubMed ID: 19762341). In epithelial cells, the TGF-β pathway normally inhibits growth and proliferation; pathogenic variants in BMPR1A and SMAD4 decrease TGF-β signaling and lead to neoplasia. BMPR1A encodes a transmembrane serine/threonine kinase receptor that binds the bone morphogenetic protein (BMP) subfamily of TGF-β ligands (Heldin et al. 1997. PubMed ID: 9393997). Approximately 140 pathogenic variations have been identified throughout the BMPR1A gene and most (~90%) are detectable by DNA sequencing (Human Gene Mutation Database). In addition to causing JPS, one BMPR1A mutation (p.Ala338Asp) has also been identified in a family with Cowden syndrome, indicating BMPR1A pathogenic variants might also define a small subset of Cowden syndrome cases (Zhou et al. 2001. PubMed ID: 11536076). SMAD4 mediates the biological effects of the transforming growth factor-β (TGF-β) superfamily of cytokines (Miyazono et al. 2009. PubMed ID: 19762341). In epithelial cells, the TGF-β pathway normally inhibits growth and proliferation; mutations in SMAD4 decrease TGF-β signaling and can lead to carcinoma.
PTEN: PTEN hamartoma tumor syndrome (PHTS) is inherited in an autosomal dominant manner. PTEN is the only known gene to be associated with the disease. In addition to PHTS, germline pathogenic variants in PTEN have been identified in 16% of patients with autism spectrum disorders (ASD) and macrocephaly, 12.5% of patients with adenomatous and hyperplastic polyps, and 5% of women with at least two different types of cancer (Zbuk and Eng. 2006. PubMed ID: 17167516; Lintas and Persico. 2009. PubMed ID: 18728070). To date, >600 pathogenic variants have been reported for the PTEN gene, and most (~95%) can be detected by DNA sequencing (Human Gene Mutation Database). The PTEN gene consists of 9 exons and encodes a dual lipid and protein phosphatase. Pathogenic variants have been reported throughout the coding region, and sequencing of all 9 exons is recommended (Eng. 2003. PubMed ID: 12938083). Five pathogenic variants have also been reported within the minimal promoter about 800 bp upstream of the start codon, and sequencing of this region is also recommended (Teresi et al. 2007. PubMed ID: 17847000).
APC: Familial adenomatous polyposis (FAP) is an autosomal dominant disorder caused by germline pathogenic variants in the adenomatous polyposis coli (APC) gene. More than 1,900 pathogenic variants have been reported in APC (Human Gene Mutation Database), and >90% are nonsense or frameshift mutations that result in a dysfunctional, truncated protein product (Nagase and Nakamura. 1993. PubMed ID: 8111410). Germline pathogenic variants are spread throughout the coding region (Béroud and Soussi. 1996. PubMed ID: 8594558). Several pathogenic variants have also been documented in the promoter, 3’ untranslated region (UTR), and deep within intron 14 (Heinimann et al. 2001. PubMed ID: 11606402). Severe FAP (more than 1,000 polyps) typically occurs in patients with pathogenic variants between codons 1250 and 1464 (Caspari et al. 1994. PubMed ID: 7906810). In contrast, patients with attenuated FAP (fewer than 100 colorectal polyps) usually have pathogenic variants at the very 5’ and 3’ ends of the gene, or in an alternatively spliced region of exon 9 (Young et al. 1998. PubMed ID: 9603437; Soravia et al. 1998. PubMed ID: 9585611). Congenital hypertrophy of retinal pigment epithelium (CHRPE) is limited to patients with pathogenic variants between codons 457 and 1444 (Caspari et al. 1995. PubMed ID: 7795585). Two pathogenic missense variants, p.Ile1307Lys and p.Glu1317Lys (commonly found in Ashkenazi Jewish populations), predispose carriers to multiple colorectal adenomas (generally less than 100) and carcinoma, but with low and variable penetrance (Frayling et al. 1998. PubMed ID: 9724771). Pathogenic variants in the exon 1B promoter of APC have also been associated with gastric adenocarcinoma and proximal polyposis of the stomach (Li et al. 2016. PubMed ID: 27087319).
STK11: Peutz-Jeghers syndrome (PJS) is caused by heterozygous germline pathogenic variants in the tumor suppressor gene STK11. STK11, also called LKB1, encodes a serine/threonine kinase that inhibits cellular proliferation by promoting cell-cycle arrest (Tiainen et al. 1999. PubMed ID: 10430928). Second-hit pathogenic variants in STK11 ultimately lead to unfettered growth and tumorigenesis. To date, ~400 unique pathogenic variants have been described throughout the STK11 gene (Human Gene Mutation Database). Most (80%) are truncating pathogenic variants (frameshift, nonsense, splice-site, or exonic deletions) that result in early protein termination (Hearle. 2006. PubMed ID: 16707622). The remaining pathogenic variants are mostly missense or in-frame deletions. Large genomic deletions in STK11 have been described.
TP53: Li-Fraumeni syndrome (LFS) is inherited in an autosomal dominant manner and caused by heterozygous germline pathogenic variants in the TP53 gene (Malkin et al. 1990. PubMed ID: 1978757; Srivastava et al. 1990. PubMed ID: 2259385). TP53 encodes the often studied cellular tumor p53 antigen (Soussi. 2010. PubMed ID: 20930848). p53 is a ubiquitously expressed DNA-binding protein that plays a major role in the regulation of cell division, DNA repair, programmed cell death, and metabolism. More than 500 pathogenic variations have been reported throughout the TP53 gene, and nearly all are detectable by DNA sequencing (Human Gene Mutation Database). Three gross deletions encompassing one or more exons of the TP53 gene have been described but these account for less than 1% of all LFS patients. The risk of developing cancer for carriers of TP53 pathogenic variants has been estimated to be ~73% for men and nearly 100% for women (Chompret et al. 2000. PubMed ID: 10864200).
CHEK2: CHEK2 encodes a protein kinase that protects the genome from ionizing radiation and genotoxic insults. To date, approximately 180 pathogenic variants have been reported throughout the CHEK2 gene, and >95% are detectable by this DNA sequencing test (Human Gene Mutation Database).
MUTYH: The MUTYH gene encodes a vital component of the base excision repair (BER) system, which protects DNA from oxidative damage and the misincorporation of adenines opposite guanines during DNA replication (Lu et al. 2006. PubMed ID: 16720376). As such, the molecular profile of colorectal adenomas and carcinomas taken from MAP patients includes G:C → T:A transversions in the adenomatous polyposis coli (APC) and K-ras tumor suppressor (KRAS) genes, among others (Lipton et al. 2003. PubMed ID: 14633673; Jones et al. 2004. PubMed ID: 15083190). To date, about 100 pathogenic variants have been reported in the MUTYH gene, nearly all (~99%) of which are single nucleotide variations, small insertions or deletions, or splice-site pathogenic variants (www.insight-group.org; Human Gene Mutation Database). While MAP occurs in patients from various ethnic groups, specific MUTYH pathogenic variants are found in different populations. In European and North American MAP populations, two pathogenic missense variants, p.Tyr179Cys and p.Gly396Asp, are most common. Both homozygous and compound heterozygous pathogenic variants contribute to the disease (Jones et al. 2002. PubMed ID: 12393807). In Asian MAP populations, common pathogenic variants include missense variant p.Arg245Cys, splice-site variant c.934-2A>G, and nonsense variant p.Glu480*; in these cases only homozygous pathogenic variants have been reported to contribute to disease (Tao. 2004. PubMed ID: 15180946; Miyaki et al. 2005. PubMed ID: 15890374). The penetrance of colorectal cancer (CRC) for biallelic carriers of MUTYH pathogenic variants is nearly 100% by the age of 60 (Farrington et al. 2005. PubMed ID: 15931596).
ATM: Ataxia-telangiectasia is an autosomal recessive disorder that is caused by pathogenic variants in the ATM gene. ATM encodes a serine protein kinase (ATM) that is involved in DNA repair via phosphorylation of downstream proteins. It senses double-stranded DNA breaks, coordinates cell-cycle checkpoints prior to repair, and recruits repair proteins to damaged DNA sites (Taylor et al. 2004. PubMed ID: 15279810). Pathogenic variants in ATM result in defective checkpoint cycling. Over 1,000 private causative variants are described with no common hot spots. In North America, most affected individuals are compound heterozygotes for two ATM pathogenic variants. However, founder ATM pathogenic variants have been observed in several populations (Gatti and Perlman. 2016. PubMed ID: 20301790).
POLD1 and POLE: Polymerase proofreading associated polyposis (PPAP) is caused by pathogenic variants in two proofreading DNA repair genes POLD1 and POLE (Chubb et al. 2015. PubMed ID: 25559809). The protein products of these genes are involved in the replication of DNA and are also involved in DNA repair. POLE synthesizes the leading strand, while POLD synthesizes Okazaki fragments of the lagging strand. Pathogenic variants in the proof-reading exonuclease domains of these proteins result in the inability to correct mismatched bases during DNA replication. Pathogenic variants in the POLD1 gene result in predisposition to both colorectal and endometrial cancer, while POLE pathogenic variants predispose to colorectal cancer (Church et al. 2013. PubMed ID: 23528559). Tumors from individuals with these pathogenic variants are microsatellite stable but acquire base substitutions (Palles et al. 2013. PubMed ID: 23263490). Reported pathogenic variants in POLD1 include mostly missense variants. Reported pathogenic variants in POLE include missense variants and small deletions (Human Gene Mutation Database).
AXIN2: Oligodontia-colorectal cancer syndrome is inherited in an autosomal dominant fashion. Lammi et al. 2004 described a large, four-generation Finnish family in which nine family members presented with oligodontia (Lammi et al. 2004. PubMed ID: 15042511). All nine were found to have a heterozygous nonsense mutation (p.Arg656*) in the AXIN2 gene, while none of the family members with normal dentition had the pathogenic variant. Importantly, six (67%) of the individuals with oligodontia also developed colorectal neoplasia by the age of 62. In addition to these germline pathogenic variants, several somatic frameshift pathogenic variants in AXIN2 have been identified in colorectal tumors, further providing support for the role of AXIN2 in the development of cancer (Liu et al. 2000. PubMed ID: 11017067).
NTHL1: NTHL1 is a base excision repair gene, which has been associated with autosomal recessive adenomatous polyposis (Weren et al. 2015. PubMed ID: 25938944). One individual with polyposis, colorectal cancer, and multiple primary tumors was reported to be compound heterozygous for biallelic NTHL1 pathogenic variants (Rivera et al. 2015. PubMed ID: 26559593).
GALNT12: The GALNT12 gene is a member of the polypeptide GalNAc-transferases, which is involved in glycosylation for multiple cellular processes. Aberrant glycosylation is involved in tumorigenesis, and GALNT12 pathogenic variants have been reported as germline and somatic variants in colorectal cancer (Guda et al. 2009. PubMed ID: 19617566; Clarke et al. 2012. PubMed ID: 22461326). Almost all reported pathogenic variants are missense amino acid changes inherited in an autosomal dominant manner (Human Gene Mutation Database).
GREM1: The GREM1 gene encodes a member of the BMP (bone morphogenic protein) antagonist family. A large duplication spanning the 3' end of the SCG5 gene and a region upstream of GREM1 has been reported to be causative for autosomal dominant hereditary mixed polyposis syndrome (Jaeger. 2012. PubMed ID: 22561515). This duplication leads to increased GREM1 expression, which in turn leads to reduced BMP pathway activity resulting in colorectal cancer.
MLH3: The MLH3 gene encodes a protein that is part of the DNA mismatch repair family. It has been shown to be associated with colorectal cancer but is considered to be a low-risk gene (Wu et al. 2001. PubMed ID: 11586295; Kraus et al. 2015. PubMed ID: 25142776; Rohlin et al. 2017. PubMed ID: 27696107). A functional study showed that its protein product forms a less efficient MMR complex with MLH1 than the more common complex of MLH1 and PMS2 (Korhonen et al. 2008. PubMed ID: 18521850).
Clinical Sensitivity - Sequencing with CNV PG-Select
The clinical sensitivity of the tested genes for hereditary colorectal cancers is provided for each syndrome.
Lynch Syndrome: Depending on the clinical criteria used to reach a diagnosis (Amsterdam or “Revised Bethesda”), 30-50% and 15-20% of Lynch patients have a detectable MLH1 and MSH2 pathogenic sequence variant, respectively (Syngal et al. 2000. PubMed ID: 10978352). A pathogenic sequence variant in MSH6 is detected in <2% of patients that meet the stringent Amsterdam I criteria but is detected in ~12% of atypical Lynch/HNPCC families (Peltomäki and Vasen. 2004. PubMed ID: 15528792). A pathogenic sequence variant in PMS2 is detected in 1-2% of Lynch patients (Peltomäki and Vasen. 2004. PubMed ID: 15528792) and ~50% of constitutional mismatch repair-deficiency patients (Wimmer and Etzler. 2008. PubMed ID: 18709565). The clinical sensitivity of EPCAM sequence variants in Lynch syndrome is unknown as no sequence variants have been reported for this disease; however, sequence variants in the EPCAM gene are known to be causative for congenital tufting enteropathy. The clinical sensitivity of EPCAM deletions is 1-3% of individuals with Lynch syndrome (Kohlmann and Gruber. 2018. PubMed ID: 20301390). Lynch syndrome is also attributed to deletions in the MLH1, MSH2, MSH6, and PMS2 genes in approximately 5%, 20%, 7%, and 20% of cases, respectively (Kohlmann and Gruber. 2018. PubMed ID: 20301390).
Hereditary Diffuse Gastric Cancer (HDGC): The clinical sensitivity of CDH1 germline pathogenic variants is 30% for HDGC families (Carneiro et al. 2007. PubMed ID: 17513507). Large deletions have been detected in the CDH1 gene in up to 4% of patients (Kaurah and Huntsman. 2014. PubMed ID: 20301318).
Juvenile Polyposis Syndrome (JPS): This test is predicted to identify a BMPR1A pathogenic variant in 11-22% and a SMAD4 pathogenic variant in 20-26% of patients diagnosed with JPS. CNV analysis is predicted to identify a BMPR1A pathogenic variant in 1-2% and a SMAD4 pathogenic variant in 2-9% of patients diagnosed with JPS (Larsen Haidle and Howe. 2017. PubMed ID: 20301642).
PTEN Hamartoma Syndrome: This test is predicted to detect causative PTEN pathogenic variants in ~80% of patients with Cowden syndrome (CS), ~65% of patients with Bannayan-Riley-Ruvalcaba syndrome (BRRS), and ~20% of patients with Proteus syndrome (PS). Large deletions are predicted to be detected in ~11% of patients with BRRS, but the sensitivity is not known for other PTEN-related disorders (Eng. 2003. PubMed ID: 12938083).
Familial Adenomatous Polyposis: This test is predicted to detect >90% of causative APC pathogenic variants (Laken et al. 1999. PubMed ID: 10051640). Gross deletions/duplications have been reported in up to 12% of APC patient samples (Jasperson et al. 2017. PubMed ID: 20301519).
Peutz-Jeghers Syndrome: Approximately 55% of patients with a positive family history and 70% of patients with no family history of Peutz-Jeghers syndrome will receive positive results through STK11 sequencing. Approximately 45% of patients with a positive family history or 21% of patients with no family history of Peutz-Jeghers syndrome will have a pathogenic variant in STK11 by deletion analysis (McGarrity et al. 2016. PubMed ID: 20301443).
Li-Fraumeni Syndrome: Sequencing the TP53 gene yields positive results for approximately 95% of patients with Li-Fraumeni syndrome. Deletions in the TP53 gene have been detected in 1% of Li-Fraumeni cases (Schneider et al. 2013. PubMed ID: 20301488).
CHEK2-related Cancers: The clinical sensitivity of CHEK2 pathogenic variants in GI cancers is unknown, but cancer risk is increased. CHEK2 pathogenic variants are also reported in hereditary breast cancer and have been reported in 4.5% of cases (Castéra et al. 2014. PubMed ID: 24549055).
MUTYH-Associated Polyposis: By definition, nearly all (~99%) MAP patients have biallelic germline pathogenic variants in MUTYH. However, pathogenic variants in MUTYH are also found in ~25% of patients initially diagnosed with familial adenomatous polyposis (FAP; Sampson et al. 2003. PubMed ID: 12853198). Clinical sensitivity for MUTYH deletions/duplications is not currently known.
Ataxia-Telangiectasia: Approximately 90% of individuals with ataxia-telangiectasia have detectable pathogenic variants via sequencing of the ATM gene, and approximately 1-2% of patients with ataxia-telangiectasia have large genomic deletions involving the ATM gene (Gatti and Perlman. 2016. PubMed ID: 20301790).
POLD1 and POLE Colorectal Cancer Predisposition: Pathogenic variants in the POLD1 and POLE genes have been observed in 0.2% and 0.3-0.6% of individuals with colorectal cancer, respectively (Chubb et al. 2015. PubMed ID: 25559809).
The clinical sensitivity of the AXIN2, GALNT12, GREM1, MLH3 and NTHL1 genes in colorectal cancer is not known.
Testing Strategy
This test is performed using Next-Gen sequencing with additional Sanger sequencing as necessary.
This panel typically provides 99.97% coverage of all coding exons of the genes plus 10 bases of flanking noncoding DNA in all available transcripts along with other non-coding regions in which pathogenic variants have been identified at PreventionGenetics or reported elsewhere. We define coverage as ≥20X NGS reads or Sanger sequencing.
Deletion and duplication detection for STK11 and PMS2 is performed using NGS, but CNVs detected in these genes are confirmed via multiplex ligation-dependent probe amplification (MLPA).
DNA analysis of the PMS2 gene is complicated due to the presence of several pseudogenes. One particular pseudogene, PMS2CL, has high sequence similarity to PMS2 exons 11 to 15 (Blount et al. 2018. PubMed ID: 29286535). Next-generation sequencing (NGS) based copy number variant (CNV) analysis can detect deletions and duplications involving exons 1 to 10 of PMS2 but has less sensitivity for exons 11 through 15. Multiplex ligation-dependent probe amplification (MLPA) can detect deletions and duplications involving PMS2 exons 1 to 15. Of note, PMS2 MLPA is included in this test.
This test also includes analysis of the inversion of exons 1-7 in MSH2, and detection of the GREM1 promoter duplication.
Indications for Test
This test is suitable for individuals with a personal or family history of hereditary colorectal cancers. This test especially aids in a differential diagnosis of similar phenotypes, rules out particular syndromes, and provides the analysis of multiple genes simultaneously. Individuals with multiple, multifocal, recurrent, and early-onset (<50 years) colorectal tumors or a family history of colorectal tumors should be assessed with this panel. This test is specifically designed for heritable germline mutations and is not appropriate for the detection of somatic mutations in tumor tissue.
This test is suitable for individuals with a personal or family history of hereditary colorectal cancers. This test especially aids in a differential diagnosis of similar phenotypes, rules out particular syndromes, and provides the analysis of multiple genes simultaneously. Individuals with multiple, multifocal, recurrent, and early-onset (<50 years) colorectal tumors or a family history of colorectal tumors should be assessed with this panel. This test is specifically designed for heritable germline mutations and is not appropriate for the detection of somatic mutations in tumor tissue.
Genes
| Official Gene Symbol | OMIM ID |
|---|---|
| APC | 611731 |
| ATM | 607585 |
| AXIN2 | 604025 |
| BMPR1A | 601299 |
| CDH1 | 192090 |
| CHEK2 | 604373 |
| EPCAM | 185535 |
| FAN1 | 613534 |
| FOCAD | 614606 |
| GALNT12 | 610290 |
| GREM1 | 603054 |
| MLH1 | 120436 |
| MLH3 | 604395 |
| MSH2 | 609309 |
| MSH6 | 600678 |
| MUTYH | 604933 |
| NTHL1 | 602656 |
| PMS2 | 600259 |
| POLD1 | 174761 |
| POLE | 174762 |
| PTEN | 601728 |
| RAB43 | 0 |
| SMAD4 | 600993 |
| STK11 | 602216 |
| TP53 | 191170 |
| Inheritance | Abbreviation |
|---|---|
| Autosomal Dominant | AD |
| Autosomal Recessive | AR |
| X-Linked | XL |
| Mitochondrial | MT |
Diseases
Related Test
| Name |
|---|
| PGxome® |
Citations
- Abud et al. 2012. PubMed ID: 23329222
- Al-Tassan et al. 2002. PubMed ID: 11818965
- Bell et al. 1999. PubMed ID: 10617473
- Béroud and Soussi. 1996. PubMed ID: 8594558
- Blount et al. 2018. PubMed ID: 29286535
- Blumenthal and Dennis. 2008. PubMed ID: 18781191
- Carneiro et al. 2008. PubMed ID: 17513507
- Caspari et al. 1994. PubMed ID: 7906810
- Caspari et al. 1995. PubMed ID: 7795585
- Castéra et al. 2014. PubMed ID: 24549055
- Chompret et al. 2000. PubMed ID: 10864200
- Chow and Macrae. 2005. PubMed ID: 16246179
- Chubb et al. 2015. PubMed ID: 25559809
- Church et al. 2013. PubMed ID: 23528559
- Clarke et al. 2012. PubMed ID: 22461326
- Concannon et al. 2008. PubMed ID: 18701470
- De Coster et al. 2009. PubMed ID: 18771513
- Díaz-Llopis and Menezo. 1988. PubMed ID: 2830869
- Eng et al. 2001. PubMed ID: 11160785
- Eng. 2003. PubMed ID: 12938083
- Esteban-Jurado et al. 2014. PubMed ID: 24587672
- Farrington et al. 2005. PubMed ID: 15931596
- Fearnhead et al. 2001. PubMed ID: 11257105
- Frayling et al. 1998. PubMed ID: 9724771
- Galiatsatos and Foulkes. 2006. PubMed ID: 16454848
- Gatti and Perlman. 2016. PubMed ID: 20301790
- Grady and Carethers. 2008. PubMed ID: 18773902
- Guda et al. 2009. PubMed ID: 19617566
- Guilford et al. 2010. PubMed ID: 20373070
- Hearle et al. 2006. PubMed ID: 16707622
- Heinimann et al. 2001. PubMed ID: 11606402
- Heldin et al. 1997. PubMed ID: 9393997
- Howe et al. 2001. PubMed ID: 11381269
- Human Gene Mutation Database (Bio-base).
- Idos and Valle. 2021. PubMed ID: 20301390
- Jaeger. 2012. PubMed ID: 22561515
- Jang and Chung. 2010. PubMed ID: 20559516
- Jasperson et al. 2017. PubMed ID: 20301519
- Jones et al. 2002. PubMed ID: 12393807
- Jones et al. 2004. PubMed ID: 15083190
- Kaurah and Huntsman. 2014. PubMed ID: 20301318
- Kempers et al. 2011. PubMed ID: 21145788
- Korhonen et al. 2008. PubMed ID: 18521850
- Kraus et al. 2015. PubMed ID: 25142776
- Laken et al. 1999. PubMed ID: 10051640
- Lammi et al. 2004. PubMed ID: 15042511
- Larsen Haidle and Howe. 2017. PubMed ID: 20301642
- Lee et al. 2001. PubMed ID: 11719428
- Li et al. 2016. PubMed ID: 27087319
- Lim et al. 2003. PubMed ID: 12865922
- Lintas and Persico. 2009. PubMed ID: 18728070
- Lipton et al. 2003. PubMed ID: 14633673
- Liu et al. 2000. PubMed ID: 11017067
- Lu et al. 2006. PubMed ID: 16720376
- Malkin et al. 1990. PubMed ID: 1978757
- Matalova et al. 2008. PubMed ID: 18573979
- McGarrity et al. 2016. PubMed ID: 20301443
- Meijers-Heijboer et al. 2003. PubMed ID: 12690581
- Miyaki et al. 2005. PubMed ID: 15890374
- Miyazono et al. 2009. PubMed ID: 19762341
- Mork et al. 2016. PubMed ID: 28004223
- Nagase and Nakamura. 1993. PubMed ID: 8111410
- Narod. 2010. PubMed ID: 20597917
- Nichols et al. 2001. PubMed ID: 11219776
- Nugent et al. 1993. PubMed ID: 8395444
- Palles et al. 2013. PubMed ID: 23263490
- Peltomäki and Vasen. 2004. PubMed ID: 15528792
- Pharoah et al. 2001. PubMed ID: 11729114
- Poulsen and Bisgaard. 2008. PubMed ID: 19506731
- Rhees et al. 2013. PubMed ID: 24114314
- Rivera et al. 2015. PubMed ID: 26559593
- Rohlin et al. 2017. PubMed ID: 27696107
- Sampson et al. 2003. PubMed ID: 12853198
- Schneider et al. 2013. PubMed ID: 20301488
- Schrader and Huntsman. 2010. PubMed ID: 20517687
- Sieber et al. 2003. PubMed ID: 12606733
- Siegel et al. 2015. PubMed ID: 25559415
- Soravia et al. 1998. PubMed ID: 9585611
- Soussi. 2010. PubMed ID: 20930848
- Srivastava et al. 1990. PubMed ID: 2259385
- Swift et al. 1991. PubMed ID: 1961222
- Syngal et al. 2000. PubMed ID: 10978352
- Tao. 2004. PubMed ID: 15180946
- Taylor et al. 2004. PubMed ID: 15279810
- Teresi et al. 2007. PubMed ID: 17847000
- Thompson et al. 2005. PubMed ID: 15928302
- Tiainen et al. 1999. PubMed ID: 10430928
- Vahteristo et al. 2002. PubMed ID: 12094328
- Varley et al. 1997. PubMed ID: 9242456
- Wagner et al. 2002. PubMed ID: 12203789
- Weren et al. 2015. PubMed ID: 25938944
- Wimmer and Etzler. 2008. PubMed ID: 18709565
- Wu et al. 2001. PubMed ID: 11586295
- www.insight-group.org
- Young et al. 1998. PubMed ID: 9603437
- Zbuk and Eng. 2007. PubMed ID: 17167516
- Zhou et al. 2001. PubMed ID: 11536076
- Zhou et al. 2001. PubMed ID: 11476841
Ordering/Specimens
Ordering Options
We offer several options when ordering sequencing tests. For more information on these options, see our Ordering Instructions page. To view available options, click on the Order Options button within the test description.
myPrevent - Online Ordering
- The test can be added to your online orders in the Summary and Pricing section.
- Once the test has been added log in to myPrevent to fill out an online requisition form.
- PGnome sequencing panels can be ordered via the myPrevent portal only at this time.
Requisition Form
- A completed requisition form must accompany all specimens.
- Billing information along with specimen and shipping instructions are within the requisition form.
- All testing must be ordered by a qualified healthcare provider.
For Requisition Forms, visit our Forms page
If ordering a Duo or Trio test, the proband and all comparator samples are required to initiate testing. If we do not receive all required samples for the test ordered within 21 days, we will convert the order to the most effective testing strategy with the samples available. Prior authorization and/or billing in place may be impacted by a change in test code.
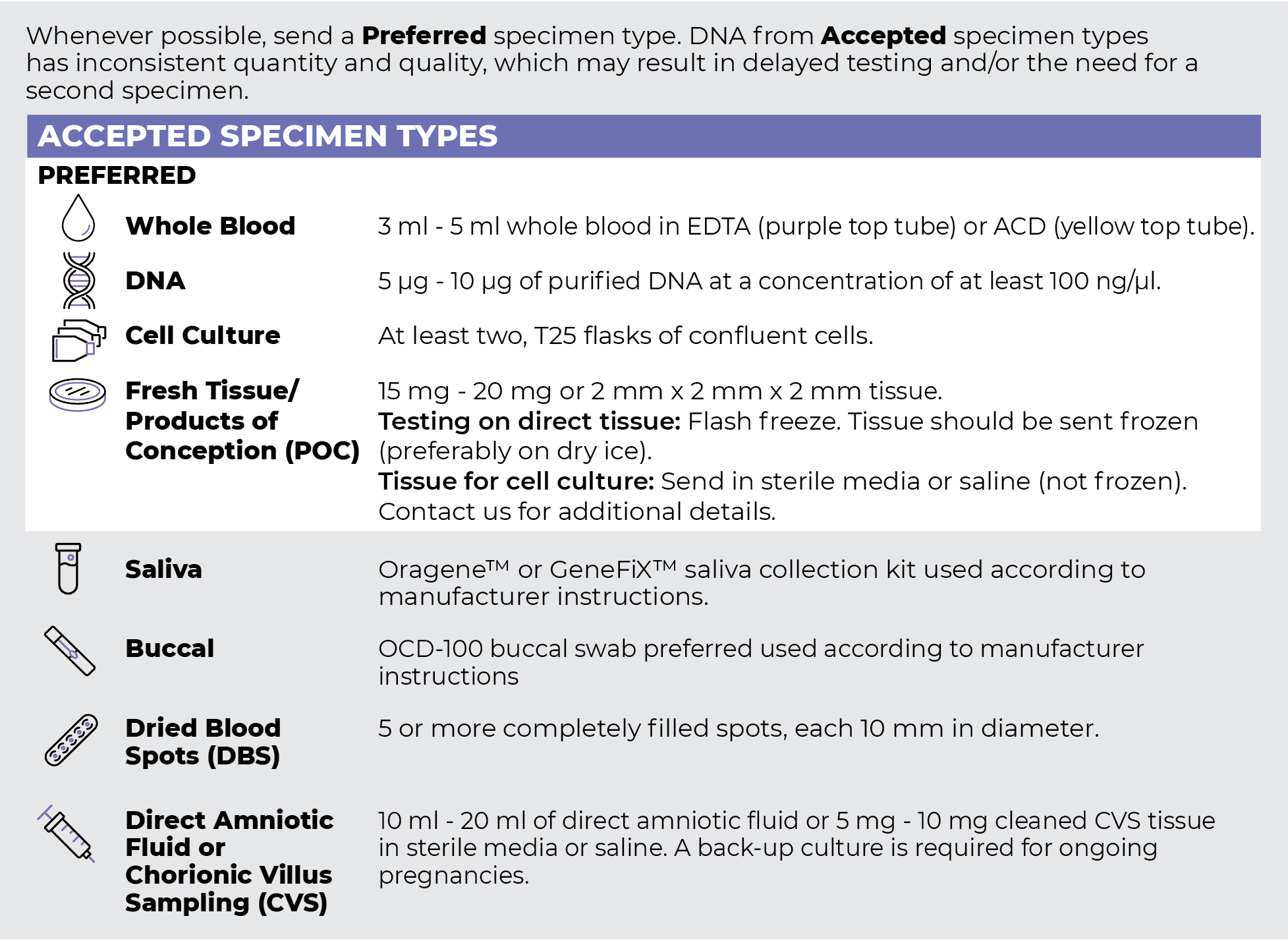
Specimen Types
Specimen Requirements and Shipping Details
ORDER OPTIONS
View Ordering Instructions1) Select Test Type
2) Select Additional Test Options
No Additional Test Options are available for this test.