



Chromosomal Instability Syndromes Panel
Summary and Pricing 
Test Method
Exome Sequencing with CNV Detection| Test Code | Test Copy Genes | Panel CPT Code | Gene CPT Codes Copy CPT Code | Base Price | |
|---|---|---|---|---|---|
| 10251 | Genes x (8) | 81479 | 81408(x1), 81479(x15) | $990 | Order Options and Pricing |
Pricing Comments
We are happy to accommodate requests for testing single genes in this panel or a subset of these genes. The price will remain the list price. If desired, free reflex testing to remaining genes on panel is available. Alternatively, a single gene or subset of genes can also be ordered via our Custom Panel tool.
An additional 25% charge will be applied to STAT orders. STAT orders are prioritized throughout the testing process.
Click here for costs to reflex to whole PGxome (if original test is on PGxome Sequencing platform).
Click here for costs to reflex to whole PGnome (if original test is on PGnome Sequencing platform).
Turnaround Time
3 weeks on average for standard orders or 2 weeks on average for STAT orders.
Please note: Once the testing process begins, an Estimated Report Date (ERD) range will be displayed in the portal. This is the most accurate prediction of when your report will be complete and may differ from the average TAT published on our website. About 85% of our tests will be reported within or before the ERD range. We will notify you of significant delays or holds which will impact the ERD. Learn more about turnaround times here.
Targeted Testing
For ordering sequencing of targeted known variants, go to our Targeted Variants page.
Clinical Features and Genetics
Clinical Features
Ataxia telangiectasia (A-T) is characterized by early onset (1-4 years) progressive cerebellar ataxia, telangiectasias of the conjunctivae, oculomotor apraxia, choreoathetosis, immunodeficiency, slurred speech, frequent infections, and an increased risk of cancers, especially leukemia and lymphoma, but other solid tumors do occur. Cancer risk is increased by 60-180 fold over population risk (Thoms et al. Experimental Dermatology 16:532–544, 2007). Other features can include premature aging (e.g. grey hair) and insulin-resistant diabetes mellitus. Individuals with A-T are usually sensitive to ionizing radiation (e.g. radiotherapy). Non-classic forms can occur with adult-onset A-T and A-T with early-onset dystonia. Unlike A-T, which is caused by homozygous mutations in the ATM gene, heterozygous carriers of an ATM mutation are at an increased risk of breast cancer (Concannon et al. Cancer Res 68:6486–91, 2008) and heart disease (Swift et al. N Engl J Med 325:1831–6, 1991). The time of onset and progression is variably between and within families of affected individuals. The prevalence of A-T has been reported to be 1:40,000-100,000 in the United States (Gatti. GeneReviews. 2010).
Ataxia-telangiectasia-like disorder (ATLD) is similar to the A-T in that patients show progressive cerebellar ataxia, however they do not show telangiectasias, and show normal immunoglobulin levels, later onset disease and a milder clinical course compared to A-T. At the cellular level, there is chromosomal instability, increased sensitivity to ionizing radiation, defective induction of stress-activated signal transduction pathways, and radioresistant DNA synthesis (Taylor et al. DNA Repair 3(8-9):1219-25, 2004).
Bloom’s Syndrome (BS) was first described in 1954 as a “congenital” skin disorder in “dwarfs” (Bloom. Am J Dis Child 88:754-758, 1954). While patients were found to have additional clinical features, such as immune deficiencies and a propensity for cancer (German et al. Science 148:506-507, 1965; German Am J Hum Genet 21-196-227, 1969), sun-sensitive facial lesions, low birth weight and stunted growth remain the most consistent features of Bloom’s Syndrome today. Symptoms of BS are related to increased mutability of proliferating somatic cells, particularly epithelial cells and lymphocytes. When grown in culture and viewed microscopically, cells of BS patients exhibit extensive “chromosome breakage”, including gaps and breaks, structural rearrangements, and telomeric associations (German and Crippa Ann Genet 9143-154, 1966). Chromosome breakage ultimately leads to excessive somatic recombination and high mutation rates (Groden and German Hum Genet 90:360-367, 1992).
Cockayne Syndrome (CS) presents with sun-sensitivity and postnatal growth failure, but does not have increased skin cancer rates. Other minor features include cachexia, neurological, psychomotoric, mental developmental delays, cataracts, retinopathy, deafness, dental caries, and characteristic facies (Thoms et al. Experimental Dermatology 16:532–544, 2007). Patients also show growth retardation, microcephaly and calcifications of the basal ganglia or elsewhere in the central nervous system. Pathologically, neurological impairment correlates with a primary demyelinization of neurons. This is in contrast with the primary neuronal degeneration found in XP patients. There are three CS forms: (1) CS Type I is the classic/moderate form usually noticed by 1-2 years of age; (2) CS Type II is the severe form noticed in the neonatal period; and (3) CS Type III is the least severe and later-onset (Laugel. GeneReviews. 2012). The prevalence of CS is 2.3 per 1,000,000 livebirths, but this is considered a conservative estimate (Kleijer et al. DNA Repair (Amst) 7:744–50, 2008).
Nijmegen breakage syndrome (NBS) is a rare autosomal recessive disease that causes microcephaly, short stature, immunodeficiency, and predisposition to cancer. Approximately half the individuals with NBS develop non-Hodgkin lymphoma or leukemia. Other cancers observed include medulloblastomas, gliomas, and rhabdomyosarcomas. Individuals with NBS are 50 times more likely to develop cancer than people without. Intellectual development is normal in 40% of patients, borderline-to-mild retardation is found in 50% of patients, and 10% of patients are moderately retarded (Kondratenko et al. Adv Exp Med Biol 601:61-7, 2007). Women with NBS often have premature ovarian insufficiency and are infertile. It is estimated that the incidence of NBS is 1 in 100,000, with a higher prevalence in specific European ancestries (i.e. Slavic population) (Concannon and Gatti. GeneReviews. 2011). Rothmund Thomson Syndrome (RTS) presents skeletal (e.g. small stature, dysplasias) and cutaneous abnormalities, with a high incidence of osteosarcomas and non-melanoma skin cancer. Individuals with RTS are photosensitive and have facial erythema and swelling in the first months of life. In addition, they present with erythema in their buttocks and extremities, while sparing the trunk, and chronic poikiloderma with hyper/hypopigmentation. Individuals with RTS also have telangiectasias, spot-like skin atrophy, and cataracts. (Thoms et al. Experimental Dermatology 16:532–544, 2007). RTS is considered a rare disorder and its prevalence is unknown (Wang and Plon. GeneReviews. 2009).
Werner Syndrome (WS) presents premature aging (e.g. greying and loss of hair) and various cancers and has been called the "adult form of progeria". Most clinical signs can be observed after 10 years of age. In addition, to premature aging there are clinical features of osteoporosis, hoarseness of voice, short stature, atherosclerosis, cataracts, hypogonadism, and diabetes. A high risk of UV-independent melanomas of the mucosae and acrolentiginous melanomas is noted. Other common cancers include soft tissue sarcomas, thyroid cancers, meningiomas, and osteosarcomas (Thoms et al. Experimental Dermatology 16:532–544, 2007). The prevalence of Werner syndrome is 1:20,000-40,000 in Japanese populations (Satoh et al. Lancet 353:1766, 1999), 1:50,000 in the Sardian population (Masala et al. Eur J Dermatol 17:213–216, 2007), and has been estimated to be 1:200,000 in the United States (Martin et al. J Am Geriatr Soc 47:1136–44, 1999).
Genetics
Cellular DNA is constantly being bombarded by endogenous damage, such as oxygen radicals, inappropriate methylation, and exogenous damage, such as chemicals, chemotherapeutics and ionizing radiation. DNA damage can be also induced during cellular processes, such as mobile element transposition, rearrangements of immune genes, and meiotic recombination (Duker. American Journal of Medical Genetics (Semin. Med. Genet.) 115:125–129, 2002). Correction of these damaging insults require a coordinated effort of genes that are involved in DNA damage sensing, checkpoint control and repair. Mutations in these genes can lead to genomic instability within syndromes leading to an elevated risk of cancers.
Ataxia-telangiectasia is an autosomal recessive disorder that is caused by homozygous mutations in the ATM gene. ATM encodes a serine protein kinase (ATM) that is involved in DNA repair via phosphorylation of downstream proteins. It senses double-stranded DNA breaks, coordinates cell-cycle checkpoints prior to repair, and recruits repair proteins to damaged DNA sites (Taylor et al. DNA Repair 3(8-9):1219-25, 2004). Mutations in ATM result in defective checkpoint cycling. Over 500 private mutations are described with no common hot spots for mutations. In North America most affected individuals are compound heterozygotes for two ATM mutations. Founder mutations for ATM mutations have been observed in several populations (Gatti. GeneReviews. 2010).
Ataxia-telangiectasia-like disorder is caused by mutations in the MRE11/MRE11A gene, which is involved in homologous recombination, telomere length maintenance, cell cycle checkpoint control and DNA double-strand break repair (Taylor et al. DNA Repair 3(8-9):1219-25, 2004). The MRE11A protein forms a complex with the RAD50 and NBS1 proteins to form the MRN complex, which is required for proper DNA repair. The serine protein kinase mutated in A-T acts upstream of the MRN complex and is responsible for MRN activation through phosphorylation. The MRN complex also has a role in ATM recruitment to DNA damage foci (Wu et al. Mol Cell 46(3):351-61, 2012) and has been shown to associate with mismatch repair proteins (Wu et al. Mol Cancer Res 9(11):1443-8, 2011). ATLD has an autosomal recessive mode of inheritance and mutations reported to date include missense, splicing, and a small deletion (Human Gene Mutation Database).
Bloom’s syndrome is an autosomal recessive disorder, caused by homozygous or compound heterozygous mutations in the BLM gene (German et al. Hum Mutat 28:743-753, 2007); more than 60 unique mutations have been identified. Most (60%) are single nucleotide changes leading to nonsense, missense or splicing mutations, while the remaining are small insertions/deletions (35%) or large deletions of multiple exons (5%). The BLM gene encodes a DNA helicase of the RecQ family. RecQ proteins are critical for maintaining the efficiency and integrity of DNA replication (Hickson Nat Rev Cancer 3:169-178, 2003); they resolve secondary structures ahead of replication forks, limit recombination to identical sequences, and assist in the replication and maintenance of telomeres (Bennett and Keck Crit Rev Biochem Mol Biol 39:79-97, 2004). In addition to these cellular functions, the BLM protein may also be important for the Mismatch Repair (MMR) pathway through its interaction with the MLH1 and MSH6 proteins (Langland et al. J Biol Chem 276:30031-30035, 2001; Pedrazzi et al. Biol Chem 384:1155-1164, 2003). Indeed, there is some evidence that heterozygous carriers of a BLM mutation have an increased risk for colorectal cancer (Gruber et al. Science 297:2013, 2002), a disease most commonly associated with heterozygous mutations in the MMR genes: MLH1, MSH2 and MSH6.
Cockayne syndrome is an autosomal recessive disorder. ERCC6 and ERCC8 appear to play an important role in the temporary removal of the stalled polymerase II to allow repair followed by the continuation of the transcription. Interestingly, nucleotide excision repair from cells of CS patients are defective in transcription couple repair (TCR), but exhibit normal global genome repair. This may help to explain the lack of cancer development, whereas both pathways are usually defective in Xeroderma Pigmentation patients who have high rates of skin cancer. Interestingly, CS cells can repair 6-4 photoproducts, unlike cells from XP patients. Finally, CS cells exhibit an increased rate of apoptosis because of TCR failure and blockage of transcription. Enhanced apoptosis of initially damaged cells may also prevent tumor cell development. (Thoms et al. Experimental Dermatology, 16:532–544, 2007). There are no reported genotype-phenotype correlations (Laugel. GeneReviews. 2012).
Nijmegen Breakage Syndrome is an autosomal recessive disease caused by mutations in the NBN gene. The protein product of NBN, Nibrin, normally associates with the MRE11A and RAD50 proteins to form the MRN complex. The MRN complex, upon DNA damage, is involved in DNA repair and cell cycle arrest via the ATM kinase; pathogenic mutations in NBN lead to faulty DNA repair and improper cell cycle control. NBS is inherited in an autosomal recessive manner, although carriers of NBN mutations may be at a higher risk of malignancies (Ciara et al. Acta Neuropathol 119(3):325-34, 2010; Steffen et al. Int J Cancer 10;111(1):67-71, 2004). Most mutations reported to date in NBN result in truncation mutations of Nibrin. The c.657_661del mutation is the most common mutation found in Eastern Europeans with NBS, accounting for more than 90% of all mutant alleles. Other mutations are private mutations which occur in one or a small number of families (Concannon and Gatti. GeneReviews. 2011).
Rothmund Thomson Syndrome is an autosomal recessive disease that is caused by mutations in the RECQL4 gene. The RECQL4 gene encodes the protein ATP-dependent DNA helicase Q4, which functions in unwinding DNA for many biological processes (e.g. replication initiation) (Sangrithi et al. Cell 121:887–98, 2005). The majority of mutations result in absent protein from truncating mutations (Siitonen et al. Eur J Hum Genet 17:151–8, 2009; Wang and Plon. GeneReviews. 2009).
Werner syndrome is an autosomal recessive disease that is caused by mutations in the WRN gene. WRN encodes an ATP-dependent helicase, belonging to the RECQ family of helicases. It is involved in unwinding DNA for processes such as DNA replication, repair, recombination, and transcription. It also has is an exonuclease and functions in the maintenance of telomeres (Rossi et al. DNA Repair (Amst) 9:331–44, 2010). The most common mutation is c.1105C>T, which is prevalent in European and Japanese populations (Matsumoto et al. Hum Genet 100:123–30, 1997; .Friedrich et al. Hum Genet 128:103–11, 2010). Founder mutations exist in other populations (Oshima et al. GeneReviews. 2012). Most mutations result in loss of protein expression via frameshift.
Clinical Sensitivity - Sequencing with CNV PGxome
Approximately 90% of individuals with Ataxia-telangiectasia have detectable mutations via sequencing of the ATM gene. Approximately 1-2% of patients have large genomic deletions involving ATM (Gatti. GeneReviews. 2010).
Clinical sensitivity is unknown for Ataxia-telangiectasia-like Disorder because it is a rare disease and relatively few patients have been reported.
In a retrospective study (German et al. Hum Mutat 28(8): 743-53, 2007), 87% of patients diagnosed with Bloom’s Syndrome were reported to have two BLM mutations. In 6% of the patients, only one mutation was found for this recessive disease, indicating the second mutation was not detectable by DNA sequencing methods.
Mutations in ERCC6 cause Cockayne syndrome complementation group type B, which account for 65% of cases. Approximately 95% of mutations will be detected by sequencing, and approximately 5% are due to gross deletions. Mutations in ERCC8 cause Cockayne syndrome complementation group type A, which accounts for 35% of cases. Approximately 80% of mutations will be detected by sequencing, and approximately 20% are due to gross deletions (Laugel. GeneReviews. 2012)
Most NBN mutations reported are based on individuals who are homozygous for the single most common Eastern European mutation, 657_661del, which can be readily detected using sequencing. In the United States, approximately 70% of individuals with NBS tested to date are homozygous for the common allele (657_661del), 15% are heterozygous for c.657_661del5 and a second unique mutation, and 15% are homozygous for a unique mutation (Concannon and Gatti. GeneReviews. 2011). Individuals homozygous for the NBN c.1089C>A mutation have features of Fanconi anemia (Gennery et al. Clin Immunol 113(2):214-9, 2004).
Sequencing of RECQL4 will detect approximately 66% of Rothmund Thomson Syndrome cases. The other 34% may be due to genetic heterogeneity although no other genes have been found. The rate of deletions/duplications is unknown (Wang and Plon. GeneReviews. 2009).
Mutations in WRN are identified by sequencing in approximately 90% of individuals with Werner syndrome (Oshima et al. GeneReviews. 2012).
Testing Strategy
This test is performed using Next-Gen sequencing with additional Sanger sequencing as necessary.
This panel typically provides 99.3% coverage of all coding exons of the genes plus 10 bases of flanking noncoding DNA in all available transcripts along with other non-coding regions in which pathogenic variants have been identified at PreventionGenetics or reported elsewhere. We define coverage as ≥20X NGS reads or Sanger sequencing. PGnome panels typically provide slightly increased coverage over the PGxome equivalent. PGnome sequencing panels have the added benefit of additional analysis and reporting of deep intronic regions (where applicable).
Dependent on the sequencing backbone selected for this testing, discounted reflex testing to any other similar backbone-based test is available (i.e., PGxome panel to whole PGxome; PGnome panel to whole PGnome).
Indications for Test
This test especially aids in a differential diagnosis of similar phenotypes, rules out particular syndromes, and provides the analysis of multiple genes simultaneously. Individuals who are suspected of any of these disorders, especially if clinical diagnosis is unclear, and individuals who have been found to be negative by mutation analysis for a single gene test are candidates. Laboratory findings that support the diagnosis include: protein levels, chromosome translocations, immunodeficiency, and radiosensitivity demonstrated by in vitro assay. This test is specifically designed for heritable germline mutations and is not appropriate for the detection of somatic mutations in tumor tissue. Individuals presenting with a Cockayne syndrome phenotype may have a mutation in the Xeroderma Pigmentosum genes ERCC2, ERCC3, or ERCC5 (Rapin et al Neurology 28;55(10):1442-9, 2000). Please see Xeroderma Pigmentosum gene testing in our test menu.
This test especially aids in a differential diagnosis of similar phenotypes, rules out particular syndromes, and provides the analysis of multiple genes simultaneously. Individuals who are suspected of any of these disorders, especially if clinical diagnosis is unclear, and individuals who have been found to be negative by mutation analysis for a single gene test are candidates. Laboratory findings that support the diagnosis include: protein levels, chromosome translocations, immunodeficiency, and radiosensitivity demonstrated by in vitro assay. This test is specifically designed for heritable germline mutations and is not appropriate for the detection of somatic mutations in tumor tissue. Individuals presenting with a Cockayne syndrome phenotype may have a mutation in the Xeroderma Pigmentosum genes ERCC2, ERCC3, or ERCC5 (Rapin et al Neurology 28;55(10):1442-9, 2000). Please see Xeroderma Pigmentosum gene testing in our test menu.
Genes
| Official Gene Symbol | OMIM ID |
|---|---|
| ATM | 607585 |
| BLM | 604610 |
| ERCC6 | 609413 |
| ERCC8 | 609412 |
| MRE11 | 600814 |
| NBN | 602667 |
| RECQL4 | 603780 |
| WRN | 604611 |
| Inheritance | Abbreviation |
|---|---|
| Autosomal Dominant | AD |
| Autosomal Recessive | AR |
| X-Linked | XL |
| Mitochondrial | MT |
Diseases
| Name | Inheritance | OMIM ID |
|---|---|---|
| Ataxia-Telangiectasia Syndrome | AR | 208900 |
| Ataxia-Telangiectasia-Like Disorder | AR | 604391 |
| Baller-Gerold Syndrome | AR | 218600 |
| Bloom Syndrome | AR | 210900 |
| Cockayne Syndrome Type I | AR | 216400 |
| Cockayne Syndrome, Type B | AR | 133540 |
| Nijmegen Breakage Syndrome | AR | 251260 |
| Rapadilino Syndrome | AR | 266280 |
| Rothmund-Thomson Syndrome | AR | 268400 |
| Werner Syndrome | AR | 277700 |
Related Test
| Name |
|---|
| PGxome® |
Citations
- Bennett RJ, Keck JL. 2004. Structure and function of RecQ DNA helicases. Crit. Rev. Biochem. Mol. Biol. 39: 79-97. PubMed ID: 15217989
- Bloom, D. 1954. Congenital telangiectatic erythema resembling lupus erythematosus in dwarfs; probably a syndrome entity. AMA Am J Dis Child 88(6): 754-8. PubMed ID: 13206391
- Ciara et al. (2010). PubMed ID: 19908051
- Concannon and Gatti. (2011). PubMed ID: 20301355
- Concannon P, Haile RW, Borresen-Dale A-L, Rosenstein BS, Gatti RA, Teraoka SN, Diep AT, Jansen L, Atencio DP, Langholz B, Capanu M, Liang X, et al. 2008. Variants in the ATM Gene Associated with a Reduced Risk of Contralateral Breast Cancer. Cancer Research 68: 6486–6491. PubMed ID: 18701470
- Duker. (2002). PubMed ID: 12407692
- Friedrich et al. (2010). PubMed ID: 20443122
- Gatti R. 2010. Ataxia-Telangiectasia. In: Pagon RA, Adam MP, Bird TD, Dolan CR, Fong C-T, and Stephens K, editors. GeneReviews™, Seattle (WA): University of Washington, Seattle. PubMed ID: 20301790
- Gennery et al. (2004). PubMed ID: 15451479
- German J, Sanz MM, Ciocci S, Ye TZ, Ellis NA. 2007. Syndrome-causing mutations of the BLM gene in persons in the Bloom’s Syndrome Registry. Human Mutation 28: 743–753. PubMed ID: 17407155
- German, J. 1969. Bloom's syndrome. I. Genetical and clinical observations in the first twenty-seven patients. Am J Hum Genet 21(2): 196-227. PubMed ID: 5770175
- German, J., et.al. 1965. Chromosomal Breakage in a Rare and Probably Genetically Determined Syndrome of Man. Science 148: 506-7. PubMed ID: 14263770
- Groden, J., German, J. (1992). PubMed ID: 1483691
- Gruber SB, Ellis NA, Scott KK, Almog R, Kolachana P, Bonner JD, Kirchhoff T, Tomsho LP, Nafa K, Pierce H, Low M, Satagopan J, et al. 2002. BLM heterozygosity and the risk of colorectal cancer. Science 297: 2013.. PubMed ID: 12242432
- Hickson ID. 2003. RecQ helicases: caretakers of the genome. Nature Reviews Cancer 3: 169–178. PubMed ID: 12612652
- Kleijer WJ, Laugel V, Berneburg M, Nardo T, Fawcett H, Gratchev A, Jaspers NGJ, Sarasin A, Stefanini M, Lehmann AR. 2008. Incidence of DNA repair deficiency disorders in western Europe: Xeroderma pigmentosum, Cockayne syndrome and trichothiodystrophy. DNA Repair 7: 744–750. PubMed ID: 18329345
- Kondratenko et al. (2007). PubMed ID: 17712992
- Langland G. 2001. The Bloom’s Syndrome Protein (BLM) Interacts with MLH1 but Is Not Required for DNA Mismatch Repair. Journal of Biological Chemistry 276: 30031–30035. PubMed ID: 11325959
- Laugel. (2012). PubMed ID: 20301516
- Martin et al. (1999). PubMed ID: 10484259
- Masala et al. (2007). PubMed ID: 17478382
- Matsumoto et al. (1997). PubMed ID: 9225981
- Oshima et al. (2012). PubMed ID: 20301687
- Pedrazzi G, Bachrati CZ, Selak N, Studer I, Petkovic M, Hickson ID, Jiricny J, Stagljar I. 2003. The Bloom’s syndrome helicase interacts directly with the human DNA mismatch repair protein hMSH6. Biol. Chem. 384: 1155–1164. PubMed ID: 12974384
- Rapin I, Lindenbaum Y, Dickson DW, Kraemer KH, Robbins JH. 2000. Cockayne syndrome and xeroderma pigmentosum. Neurology 55: 1442–1449. PubMed ID: 11185579
- Rossi et al. (2010). PubMed ID: 20075015
- Sangrithi et al. (2005). PubMed ID: 15960976
- Satoh et al. (1999). PubMed ID: 10347997
- Siitonen et al. (2009). PubMed ID: 18716613
- Steffen et al. (2004). PubMed ID: 15185344
- Swift M, Morrell D, Massey RB, Chase CL. 1991. Incidence of cancer in 161 families affected by ataxia-telangiectasia. N. Engl. J. Med. 325: 1831–1836. PubMed ID: 1961222
- Taylor et al. (2004). PubMed ID: 15279810
- Thoms et al. (2007). PubMed ID: 17518994
- Wang and Plon. (2009). PubMed ID: 20301415
- Wu et al. (2011). PubMed ID: 21849470
Ordering/Specimens
Ordering Options
We offer several options when ordering sequencing tests. For more information on these options, see our Ordering Instructions page. To view available options, click on the Order Options button within the test description.
myPrevent - Online Ordering
- The test can be added to your online orders in the Summary and Pricing section.
- Once the test has been added log in to myPrevent to fill out an online requisition form.
- PGnome sequencing panels can be ordered via the myPrevent portal only at this time.
Requisition Form
- A completed requisition form must accompany all specimens.
- Billing information along with specimen and shipping instructions are within the requisition form.
- All testing must be ordered by a qualified healthcare provider.
For Requisition Forms, visit our Forms page
If ordering a Duo or Trio test, the proband and all comparator samples are required to initiate testing. If we do not receive all required samples for the test ordered within 21 days, we will convert the order to the most effective testing strategy with the samples available. Prior authorization and/or billing in place may be impacted by a change in test code.
Specimen Types
Specimen Requirements and Shipping Details
PGxome (Exome) Sequencing Panel
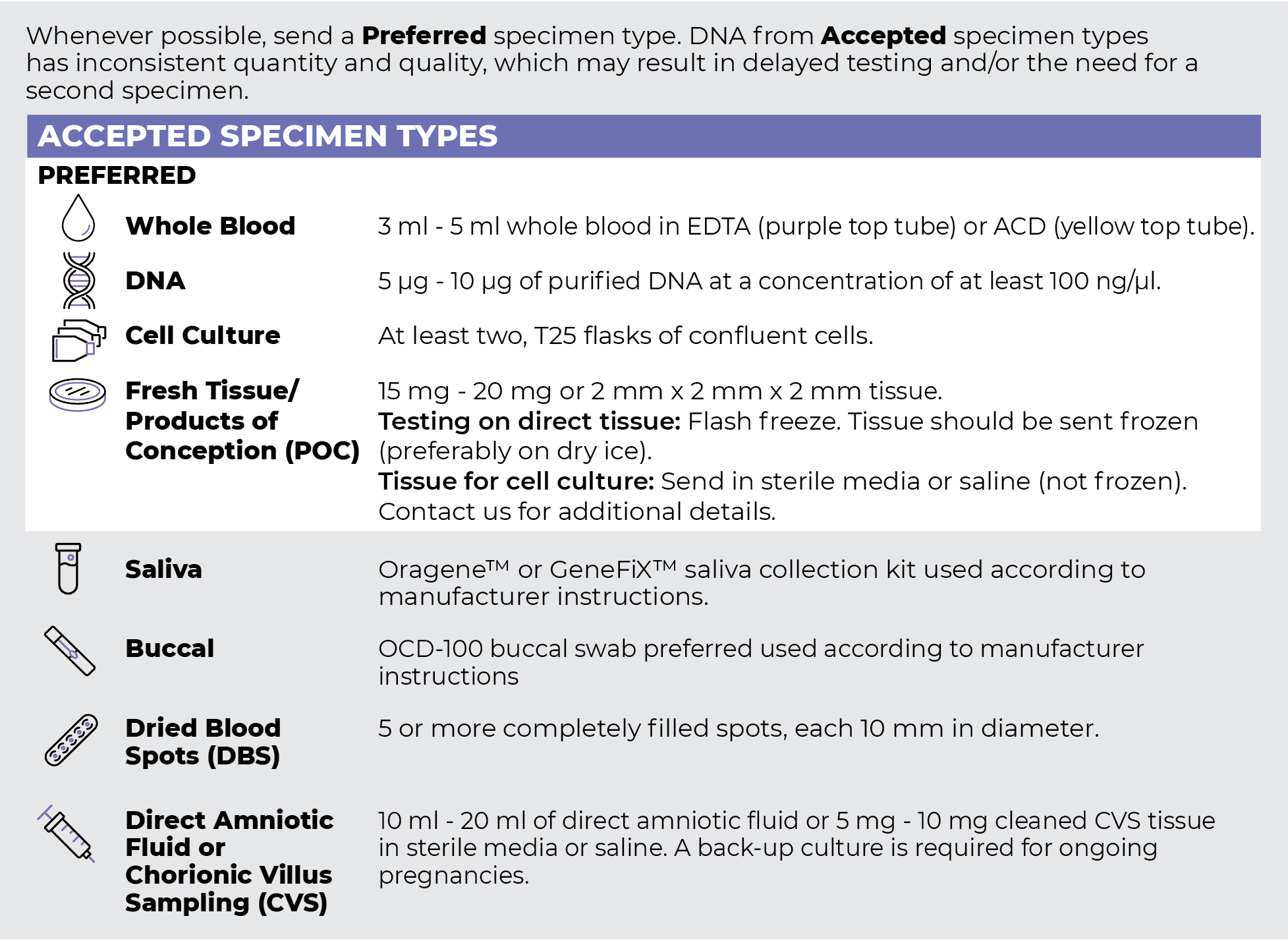
PGnome (Genome) Sequencing Panel

ORDER OPTIONS
View Ordering Instructions1) Select Test Type
2) Select Additional Test Options
No Additional Test Options are available for this test.