



Craniosynostosis and Related Disorders Panel
Summary and Pricing 
Test Method
Exome Sequencing with CNV Detection| Test Code | Test Copy Genes | Panel CPT Code | Gene CPT Codes Copy CPT Code | Base Price | |
|---|---|---|---|---|---|
| 10197 | Genes x (5) | 81479 | 81403(x1), 81404(x1), 81405(x1), 81479(x7) | $990 | Order Options and Pricing |
Pricing Comments
We are happy to accommodate requests for testing single genes in this panel or a subset of these genes. The price will remain the list price. If desired, free reflex testing to remaining genes on panel is available. Alternatively, a single gene or subset of genes can also be ordered via our Custom Panel tool.
An additional 25% charge will be applied to STAT orders. STAT orders are prioritized throughout the testing process.
Click here for costs to reflex to whole PGxome (if original test is on PGxome Sequencing platform).
Click here for costs to reflex to whole PGnome (if original test is on PGnome Sequencing platform).
Turnaround Time
3 weeks on average for standard orders or 2 weeks on average for STAT orders.
Please note: Once the testing process begins, an Estimated Report Date (ERD) range will be displayed in the portal. This is the most accurate prediction of when your report will be complete and may differ from the average TAT published on our website. About 85% of our tests will be reported within or before the ERD range. We will notify you of significant delays or holds which will impact the ERD. Learn more about turnaround times here.
Targeted Testing
For ordering sequencing of targeted known variants, go to our Targeted Variants page.
Clinical Features and Genetics
Clinical Features
Craniosynostosis is a primary abnormality of premature fusion of the cranial sutures causing skull deformity, which can occur as non-syndromic or syndromic craniosynostosis with an approximate prevalence of 1 in 2,000 to 2,500 live births worldwide. Craniosynostosis related disorders include, but are not limited to the following disorders:
Achondroplasia is characterized by abnormal bone growth that results in short stature with disproportionately short arms and legs, a large head, and characteristic facial features with frontal bossing and mid-face retrusion (Pauli 2012).
Skeletal features of Hypochondroplasia are similar to achondroplasia, but are usually milder and show failure to grow as toddlers or school-age (Bober et al. 2013).
Thanatophoric dysplasia is a perinatal lethal short-limb dwarfism syndrome, which is divided into two subtypes: type I is characterized by micromelia with bowed femurs and usually without cloverleaf skull deformity; and type II is characterized by micromelia with straight femurs, and moderate-to-severe cloverleaf skull deformity (Karczeski and Cutting 2013). Other features of thanatophoric dysplasia are: short ribs, narrow thorax, macrocephaly, distinctive facial features, brachydactyly, hypotonia, and redundant skin folds along the limbs. Most affected infants die of respiratory insufficiency shortly after birth. Rare long-term survivors have been reported.
Crouzon syndrome is characterized by hypertelorism, exophthalmos and external strabismus, parrot-beaked nose, short upper lip, hypoplastic maxilla, and a relative mandibular prognathism (Vulliamy et al. 1966).
Apert syndrome is characterized by craniosynostosis, midface hypoplasia, and syndactyly of the hands and feet with a tendency to fusion of bony structures (Glaser et al. 2003).
CATSHL syndrome is characterized by camptodactyly, tall stature and hearing loss; some less common features include kyphoscoliosis, mental retardation, learning disabilities, and microcephaly (Toydemir et al. 2006).
Muenke syndrome is characterized by uni- or bicoronal synostosis, macrocephaly, midfacial hypoplasia, and developmental delay, other features include temporal bossing; widely spaced eyes, ptosis or proptosis (usually mild); midface retrusion; and highly arched palate or cleft lip and palate. Strabismus is common (Agochukwu et al. 2014). Trigonocephaly is a disorder caused by premature closure of the metopic sutures (Azimi et al. 2002).
Pfeiffer syndrome is characterized by coronal craniosynostosis, midface hypoplasia, and broad and medially deviated thumbs and great toes (Robin et al. 2011). Osteoglophonic dysplasia is characterized by rhizomelic dwarfism with depression of the nasal bridge, frontal bossing, and prognathism (Farrow 2006).
Jackson-Weiss syndrome is characterized by premature fusion of the cranial sutures and radiographic anomalies of the feet, and normal hands (Heike et al. 2001; Cohen et al. 2001).
Hartsfield syndrome is characterized by holoprosencephaly, ectrodactyly, with or without cleft/lip palate. Some patients may also present profound mental retardation and midline and limb defects (Vilain et al. 2009).
LADD syndrome (also called Levy-Hollister Syndrome) is the short name of Lacrimoauriculodentodigital syndrome, which is featured by abnormalities of the nasal lacrimal ducts, cup-shaped pinnas with mixed hearing deficit, small and peg-shaped lateral maxillary incisors and mild enamel dysplasia and fifth finger clinodactyly, duplication of the distal phalanx of the thumb, triphalangeal thumb, and syndactyly (Thompson et al. 1985).
Saethre-Chotzen syndrome is a craniosynostosis with low frontal hairline, facial asymmetry, brachydactyly, fifth finger clinodactyly, partial syndactyly, and vertebral column defects (Reardon and Winter 1994).
Bent bone dysplasia syndrome is characterized by poor mineralization of the calvarium, craniosynostosis, dysmorphic facial features, prenatal teeth, hypoplastic pubis and clavicles, osteopenia, and bent long bones (Merrill et al. 2012).
Genetics
Hartsfield syndrome, caused by pathogenic variants in FGFR1, is inherited in both an autosomal dominant and recessive manner (Simonis et al. 2013), while FGFR1-related Pfeiffer syndrome, Trigonocephaly, Osteoglophonic dysplasia, Jackson-Weiss syndrome and Kallmann syndrome are inherited in an autosomal dominant manner. FGFR1 protein encoded by FGFR1 is a growth factor receptor and a member of the FGFR family. Like all of the FGFRs, FGFR1 is a membrane-spanning tyrosine kinase receptor with an extracellular ligand-binding domain consisting of three immunoglobulin subdomains, a transmembrane domain, and a split intracellular tyrosine kinase domain (Green et al. 1996). To date, more than 150 unique causative variants have been reported in the FGFR1 gene. These variants are: missense (70%), nonsense (7%), splicing (7%), small insertion/deletions (13%) and only 4 gross deletions and genomic complex rearrangements (Human Gene Mutation Database). The majority (~90%) of reported pathogenic variants in the FGFR1 gene are found in patients affected with Kallmann or Kallmann-related disorders (Human Gene Mutation Database). Six of the missense variants were found in patients affected with Hartsfield syndrome. Only a few disease causing FGFR1 variants were reported in other FGFR1-related disorders, such as the c.755C>G (p.Pro252Arg) variant reported to cause Pfeiffer syndrome.
FGFR2-related disorders are inherited in an autosomal dominant manner. FGFR2 protein encoded by FGFR2 is a growth factor receptor, a member of the FGFR family. Like all of the FGFRs, FGFR2 is a membrane-spanning tyrosine kinase receptor with an extracellular ligand-binding domain consisting of three immunoglobulin subdomains, a transmembrane domain, and a split intracellular tyrosine kinase domain (Green et al. 1996). To date, more than 100 unique causative variants have been reported in the FGFR2 gene. These variants are: missense (66%), splicing (13%), small insertion/deletions (18%) and only 5 gross deletions and genomic complex rearrangements (Human Gene Mutation Database; Bochukova et al. 2009). Some genotype-phenotype correlations and recurrent pathogenic variants in FGFR2 have been documented, such as p.Ser252Trp and p.Pro253Arg which are responsible for 98% of Apert syndrome cases (Bochukova et al. 2009), while p.Cys342Tyr and p.Cys342Arg are often seen in Pfeiffer or Crouzon syndrome (Rutland et al. 1995; Mulvihill et al. 1995). For Crouzon syndrome or Pfeiffer syndrome, ~80% of FGFR2 pathogenic variants are located in exons 8 and 10, and ~10% of them are in exons 3, 5, 11, 14, 15, 16, and 17 (Robin et al. 2011). FGFR2 pathogenic variants were found in 100% patients (227 patients) with Apert syndrome: 223/227 with point mutations and 4/227 with an Alu insertion or exon deletion (Bochukova et al. 2009).
All FGFR3-related disorders are inherited in an autosomal dominant manner, and the majority of cases result from de novo pathogenic variants caused by gain of functional variants. An exception is FGFR3-related CATSHL syndrome,which can be inherited either in an autosomal dominant manner or autosomal recessive manner through loss of function variants in the FGFR3 gene. The FGFR3 gene encodes fibroblast growth factor receptor-3, a member of the FGFR family. Like all of the FGFRs, FGFR3 is a membrane-spanning tyrosine kinase receptor with an extracellular ligand-binding domain consisting of three immunoglobulin subdomains, a transmembrane domain, and a split intracellular tyrosine kinase domain (Green et al. 1996). Some genotype-phenotype correlations have been well established. For example, most cases of achondroplasia are caused by one of two variants (c.1138G>A, p.Gly380Arg /c.1138G>C, p.Gly380Arg) in exon 10 (Shiang et al. 1994; Bellus et al. 1995; Deng et al. 1996). ~70% of Hypochondroplasia cases are caused by two variants (c.1620C>A, p.Asn540Lys and c.1620C>G, p.Asn540Lys) in exon 13 (Bellus et al. 1995; Prinos et al. 1995). Thanatophoric dysplasia type II is caused by the pathogenic variant c.1948A>G (p.Lys650Glu) in exon 15 (Bellus et al. 2000); and Muenke syndrome is caused by the c.749C>G, p.Pro250Arg variant in exon 7. Crouzon syndrome with acanthosis nigricans is caused by the pathogenic variant c.1172C>A, p. Ala391Glu in exon 10. In addition, two variants were found in two large families with CATSHL syndrome: heterozygous c.1862G>A, p.Arg621His in a family with autosomal dominant inheritance of CATSHL syndrome (Toydemir et al. 2006) and homozygous c.1637C>A, p.Thr546Lys in a family with autosomal recessive inheritance of CATSHL syndrome, respectively (Makrythanasis et al. 2014).
TCF12-related Craniosynostosis is inherited in an autosomal dominant manner. The TCF12 protein coded by the TCF12 gene is a member of the class A basic helix-loop-helix family, a partner of TWIST1 protein, which may be involved in the initiation of neuronal differentiation. Approximately 40 TCF12 pathogenic variants have been reported. They are: missense (7%), nonsense: (32%), splicing (17%), small deletion/insertions (41%), and one unbalanced translocation involving the TFC12 gene (Sharma et al. 2013; di Rocco et al. 2014; Le Tanno et al. 2014; Human Gene Mutation Database).
Saethre-Chotzen syndrome is inherited in an autosomal dominant manner. The TWIST1 protein coded by the TWIST1 gene is a transcription factor in the helix-loop-helix family which regulates embryonic development of many organs. Currently, ~170 unique TWIST1 pathogenic variants have been reported. They are: missense (34%), nonsense: (15%), small deletion/insertions (32%), large deletions (11%), and translocation/inversions (6%) (Johnson et al. 1998; Kress et al. 2006; Human Gene Mutation Database).
See individual gene test descriptions for additional information on molecular biology of gene products.
Clinical Sensitivity - Sequencing with CNV PGxome
~61% (111/182) of 182 Spanish craniosynostosis probands harbor a pathogenic variant in one of the FGFR2, FGFR3, TWIST1 and TCF12 genes. Pathogenic variants in FGFR2, FGFR3, TWIST and TCF12 account for 36%, 16%, 8% and 3% of pathogenic variants identified in this study, respectively (Paumard-Hernández et al. 2014).
TCF12 explains 32% and 10% of patients affected with bilateral and unilateral Craniosynostosis, respectively (Sharma et al. 2013).
One study reported that six unique FGFR1 pathogenic missense variants were found in seven unrelated patients affected with Hartsfield syndrome (Simonis et al. 2013). FGFR1 explains 5% of Pfeiffer syndrome type 1 cases (Robin et al. 2011).
Only five documented pathogenic variants in FGFR2 are large deletions/insertions (Human Gene Mutation Database; Bochukova et al. 2009). To date, no gross deletions or duplications have been reported in FGFR3 (Human Gene Mutation Database).
Testing Strategy
This test is performed using Next-Gen sequencing with additional Sanger sequencing as necessary.
This panel provides 100% coverage of all coding exons of the genes plus 10 bases of flanking noncoding DNA in all available transcripts along with other non-coding regions in which pathogenic variants have been identified at PreventionGenetics or reported elsewhere. We define coverage as ≥20X NGS reads or Sanger sequencing. PGnome panels typically provide slightly increased coverage over the PGxome equivalent. PGnome sequencing panels have the added benefit of additional analysis and reporting of deep intronic regions (where applicable).
Dependent on the sequencing backbone selected for this testing, discounted reflex testing to any other similar backbone-based test is available (i.e., PGxome panel to whole PGxome; PGnome panel to whole PGnome).
Indications for Test
Candidates for this test are patients with clinical and radiologic features consistent Craniosynostosis and Related disorders. This test especially aids in a differential diagnosis of similar phenotypes, rules out particular syndromes, and provides the analysis of multiple genes simultaneously. Individuals who are suspected of any of these disorders, especially if clinical diagnosis is unclear, and individuals who have been found to be negative by mutation analysis for single gene tests are also candidates.
Candidates for this test are patients with clinical and radiologic features consistent Craniosynostosis and Related disorders. This test especially aids in a differential diagnosis of similar phenotypes, rules out particular syndromes, and provides the analysis of multiple genes simultaneously. Individuals who are suspected of any of these disorders, especially if clinical diagnosis is unclear, and individuals who have been found to be negative by mutation analysis for single gene tests are also candidates.
Genes
| Official Gene Symbol | OMIM ID |
|---|---|
| FGFR1 | 136350 |
| FGFR2 | 176943 |
| FGFR3 | 134934 |
| TCF12 | 600480 |
| TWIST1 | 601622 |
| Inheritance | Abbreviation |
|---|---|
| Autosomal Dominant | AD |
| Autosomal Recessive | AR |
| X-Linked | XL |
| Mitochondrial | MT |
Diseases
Related Test
| Name |
|---|
| PGxome® |
Citations
- Agochukwu NB, Doherty ES, Muenke M. 2014. Muenke Syndrome. In: Pagon RA, Adam MP, Ardinger HH, Wallace SE, Amemiya A, Bean LJ, Bird TD, Dolan CR, Fong C-T, Smith RJ, and Stephens K, editors. GeneReviews(®), Seattle (WA): University of Washington, Seattle. PubMed ID: 20301588
- Azimi C, Kennedy SJ, Chitayat D, Chakraborty P, Clarke JTR, Forrest C, Teebi AS. 2002. Clinical and genetic aspects of trigonocephaly: A study of 25 cases. American Journal of Medical Genetics Part A 117A: 127–135. PubMed ID: 12567409
- Bellus GA, Hefferon TW, Ortiz de Luna RI, Hecht JT, Horton WA, Machado M, Kaitila I, McIntosh I, Francomano CA. 1995. Achondroplasia is defined by recurrent G380R mutations of FGFR3. Am. J. Hum. Genet. 56: 368-373. PubMed ID: 7847369
- Bellus GA, McIntosh I, Smith EA, Aylsworth AS, Kaitila I, Horton WA, Greenhaw GA, Hecht JT, Francomano CA. 1995. A recurrent mutation in the tyrosine kinase domain of fibroblast growth factor receptor 3 causes hypochondroplasia. Nat. Genet. 10: 357–359. PubMed ID: 7670477
- Bellus GA, Spector EB, Speiser PW, Weaver CA, Garber AT, Bryke CR, Israel J, Rosengren SS, Webster MK, Donoghue DJ, Francomano CA. 2000. Distinct missense mutations of the FGFR3 lys650 codon modulate receptor kinase activation and the severity of the skeletal dysplasia phenotype. Am. J. Hum. Genet. 67: 1411–1421. PubMed ID: 11055896
- Bober MB, Bellus GA, Nikkel SM, Tiller GE. 2013. Hypochondroplasia. In: Pagon RA, Adam MP, Ardinger HH, Wallace SE, Amemiya A, Bean LJ, Bird TD, Dolan CR, Fong C-T, Smith RJ, and Stephens K, editors. GeneReviews(®), Seattle (WA): University of Washington, Seattle. PubMed ID: 20301650
- Bochukova EG, Roscioli T, Hedges DJ, Taylor IB, Johnson D, David DJ, Deininger PL, Wilkie AOM. 2009. Rare mutations of FGFR2 causing apert syndrome: identification of the first partial gene deletion, and an Alu element insertion from a new subfamily. Hum. Mutat. 30: 204–211. PubMed ID: 18726952
- Cohen MM. 2001. Jackson-Weiss syndrome. American journal of medical genetics 100: 325–329. PubMed ID: 11343324
- Deng C, Wynshaw-Boris A, Zhou F, Kuo A, Leder P. 1996. Fibroblast growth factor receptor 3 is a negative regulator of bone growth. Cell 84: 911-921. PubMed ID: 8601314
- di Rocco F, Baujat G, Arnaud E, Rénier D, Laplanche J-L, Daire VC, Collet C. 2014. Clinical spectrum and outcomes in families with coronal synostosis and TCF12 mutations. European Journal of Human Genetics 22: 1413–1416. PubMed ID: 24736737
- Farrow EG, Davis SI, Mooney SD, Beighton P, Mascarenhas L, Gutierrez YR, Pitukcheewanont P, White KE. 2006. Extended mutational analyses of FGFR1 in osteoglophonic dysplasia. American Journal of Medical Genetics Part A 140A: 537–539. PubMed ID: 16470795
- Glaser RL, Broman KW, Schulman RL, Eskenazi B, Wyrobek AJ, Jabs EW. 2003. The Paternal-Age Effect in Apert Syndrome Is Due, in Part, to the Increased Frequency of Mutations in Sperm. Am J Hum Genet 73: 939–947. PubMed ID: 12900791
- Green PJ, Walsh FS, Doherty P. 1996. Promiscuity of fibroblast growth factor receptors. Bioessays 18: 639–646. PubMed ID: 8760337
- Heike C, Seto M, Hing A, Palidin A, Hu FZ, Preston RA, Ehrlich GD, Cunningham M. 2001. Century of Jackson-Weiss syndrome: Further definition of clinical and radiographic findings in “lost” descendants of the original kindred. American journal of medical genetics 100: 315–324. PubMed ID: 11343323
- Human Gene Mutation Database (Bio-base).
- Johnson D, Horsley SW, Moloney DM, Oldridge M, Twigg SR, Walsh S, Barrow M, Njølstad PR, Kunz J, Ashworth GJ, Wall SA, Kearney L, Wilkie AO. 1998. A comprehensive screen for TWIST mutations in patients with craniosynostosis identifies a new microdeletion syndrome of chromosome band 7p21.1. Am J Hum Genet 63: 1282-1293. PubMed ID: 9792856
- Karczeski B, Cutting GR. 2013. Thanatophoric Dysplasia. In: Pagon RA, Adam MP, Ardinger HH, Wallace SE, Amemiya A, Bean LJ, Bird TD, Dolan CR, Fong C-T, Smith RJ, and Stephens K, editors. GeneReviews(®), Seattle (WA): University of Washington, Seattle. PubMed ID: 20301540
- Kress W, Schropp C, Lieb G, Petersen B, Büsse-Ratzka M, Kunz J, Reinhart E, Schäfer W-D, Sold J, Hoppe F, Pahnke J, Trusen A, Sörensen N, Krauss J. 2006. Saethre-Chotzen syndrome caused by TWIST 1 gene mutations: functional differentiation from Muenke coronal synostosis syndrome. Eur. J. Hum. Genet. 14: 39-48. PubMed ID: 16251895
- Le Tanno P, Poreau B, Devillard F, Vieville G, Amblard F, Jouk P-S, Satre V, Coutton C. 2014. Maternal complex chromosomal rearrangement leads to TCF12 microdeletion in a patient presenting with coronal craniosynostosis and intellectual disability. Am. J. Med. Genet. 164: 1530–1536. PubMed ID: 24648389
- Makrythanasis P, Temtamy S, Aglan MS, Otaify GA, Hamamy H, Antonarakis SE. 2014. A Novel Homozygous Mutation in FGFR3 Causes Tall Stature, Severe Lateral Tibial Deviation, Scoliosis, Hearing Impairment, Camptodactyly, and Arachnodactyly. Human Mutation 35: 959-963. PubMed ID: 24864036
- Merrill AE, Sarukhanov A, Krejci P, Idoni B, Camacho N, Estrada KD, Lyons KM, Deixler H, Robinson H, Chitayat D, Curry CJ, Lachman RS, Wilcox WR, Krakow D. 2012. Bent Bone Dysplasia-FGFR2 type, a Distinct Skeletal Disorder, Has Deficient Canonical FGF Signaling. Am J Hum Genet 90: 550–557. PubMed ID: 22387015
- Mulvihill JJ. 1995. Craniofacial syndromes: no such thing as a single gene disease. Nat. Genet. 9: 101–103. PubMed ID: 7719329
- Pauli RM. 2012. Achondroplasia. In: Pagon RA, Adam MP, Ardinger HH, Wallace SE, Amemiya A, Bean LJ, Bird TD, Dolan CR, Fong C-T, Smith RJ, and Stephens K, editors. GeneReviews(®), Seattle (WA): University of Washington, Seattle. PubMed ID: 20301331
- Paumard-Hernández B, Berges-Soria J, Barroso E, Rivera-Pedroza CI, Pérez-Carrizosa V, Benito-Sanz S, López-Messa E, Santos F, García-Recuero II, Romance A, Ballesta-Martínez JM, López-González V, Campos-Barros A, Cruz J5, Guillén-Navarro E, Sánchez Del Pozo J, Lapunzina P, García-Miñaur S, Heath KE. 2014. Expanding the mutation spectrum in 182 Spanish probands with craniosynostosis: identification and characterization of novel TCF12 variants. European Journal of Human Genetics. PubMed ID: 25271085
- Prinos P, Costa T, Sommer A, Kilpatrick MW, Tsipouras P. 1995. A common FGFR3 gene mutation in hypochondroplasia. Human molecular genetics 4: 2097–2101. PubMed ID: 8589686
- Reardon W, Winter RM. 1994. Saethre-Chotzen syndrome. J Med Genet 31: 393–396. PubMed ID: 8064818
- Robin NH, Falk MJ, Haldeman-Englert CR. 2011. FGFR-Related Craniosynostosis Syndromes. In: Pagon RA, Adam MP, Bird TD, Dolan CR, Fong C-T, and Stephens K, editors. GeneReviews™, Seattle (WA): University of Washington, Seattle. PubMed ID: 20301628
- Rutland P, Pulleyn LJ, Reardon W, Baraitser M, Hayward R, Jones B, Malcolm S, Winter RM, Oldridge M, Slaney SF. 1995. Identical mutations in the FGFR2 gene cause both Pfeiffer and Crouzon syndrome phenotypes. Nat. Genet. 9: 173–176. PubMed ID: 7719345
- Sharma VP, Fenwick AL, Brockop MS, McGowan SJ, Goos JAC, Hoogeboom AJM, Brady AF, Jeelani NO, Lynch SA, Mulliken JB, Murray DJ, Phipps JM, Sweeney E, Tomkins SE, Wilson LC, Bennett S, Cornall RJ, Broxholme J, Kanapin A; 500 Whole-Genome Sequences (WGS500) Consortium, Johnson D, Wall SA, van der Spek PJ, Mathijssen IM, Maxson RE, Twigg SR, Wilkie AO. 2013. Mutations in TCF12, encoding a basic helix-loop-helix partner of TWIST1, are a frequent cause of coronal craniosynostosis. Nat Genet 45: 304-307. PubMed ID: 23354436
- Shiang R, Thompson LM, Zhu YZ, Church DM, Fielder TJ, Bocian M, Winokur ST, Wasmuth JJ. 1994. Mutations in the transmembrane domain of FGFR3 cause the most common genetic form of dwarfism, achondroplasia. Cell 78: 335–342. PubMed ID: 7913883
- Simonis N, Migeotte I, Lambert N, Perazzolo C, Silva DC de, Dimitrov B, Heinrichs C, Janssens S, Kerr B, Mortier G, Vliet G Van, Lepage P, Casimir G, Abramowicz M, Smits G, Vilain C. 2013. FGFR1 mutations cause Hartsfield syndrome, the unique association of holoprosencephaly and ectrodactyly. Journal of Medical Genetics 50: 585–592. PubMed ID: 23812909
- Thompson E, Pembrey M, Graham JM. 1985. Phenotypic variation in LADD syndrome. J. Med. Genet. 22: 382-385. PubMed ID: 4078868
- Toydemir RM, Brassington AE, Bayrak-Toydemir P, Krakowiak PA, Jorde LB, Whitby FG, Longo N, Viskochil DH, Carey JC, Bamshad MJ. 2006. A novel mutation in FGFR3 causes camptodactyly, tall stature, and hearing loss (CATSHL) syndrome. Am. J. Hum. Genet. 79: 935-941. PubMed ID: 17033969
- Vilain C, Mortier G, Vliet G Van, Dubourg C, Heinrichs C, Silva D de, Verloes A, Baumann C. 2009. Hartsfield holoprosencephaly-ectrodactyly syndrome in five male patients: Further delineation and review. American Journal of Medical Genetics Part A 149A: 1476–1481. PubMed ID: 19504604
- Vulliamy DG, Normandale PA. 1966. Cranio-facial Dysostosis in a Dorset Family. Arch Dis Child 41: 375–382. PubMed ID: 21032436
Ordering/Specimens
Ordering Options
We offer several options when ordering sequencing tests. For more information on these options, see our Ordering Instructions page. To view available options, click on the Order Options button within the test description.
myPrevent - Online Ordering
- The test can be added to your online orders in the Summary and Pricing section.
- Once the test has been added log in to myPrevent to fill out an online requisition form.
- PGnome sequencing panels can be ordered via the myPrevent portal only at this time.
Requisition Form
- A completed requisition form must accompany all specimens.
- Billing information along with specimen and shipping instructions are within the requisition form.
- All testing must be ordered by a qualified healthcare provider.
For Requisition Forms, visit our Forms page
If ordering a Duo or Trio test, the proband and all comparator samples are required to initiate testing. If we do not receive all required samples for the test ordered within 21 days, we will convert the order to the most effective testing strategy with the samples available. Prior authorization and/or billing in place may be impacted by a change in test code.
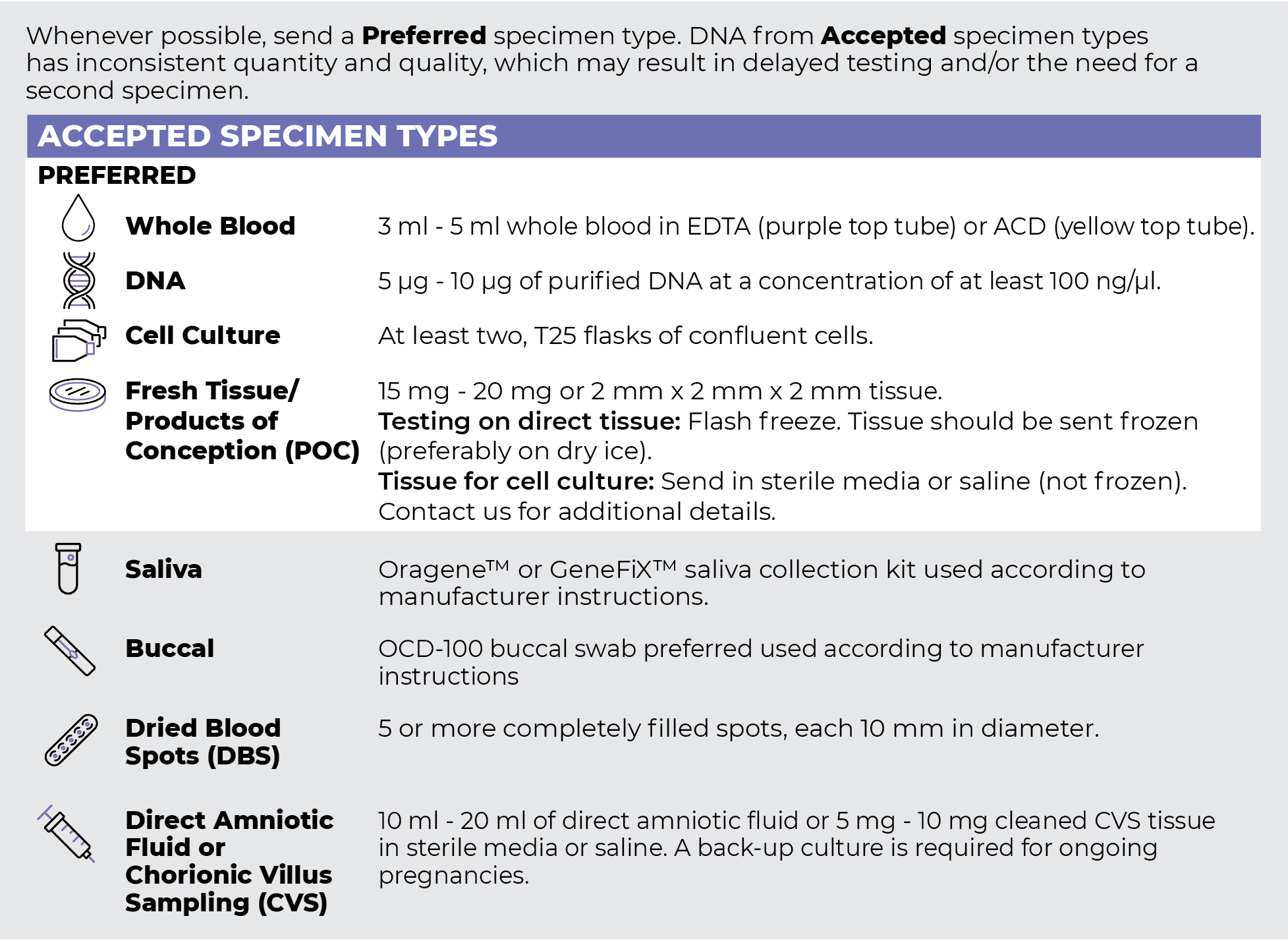
Specimen Types
Specimen Requirements and Shipping Details
PGxome (Exome) Sequencing Panel
PGnome (Genome) Sequencing Panel

ORDER OPTIONS
View Ordering Instructions1) Select Test Type
2) Select Additional Test Options
No Additional Test Options are available for this test.